点击蓝字 关注我们
家庭中宠物犬与宠物犬主人耐药基因的共存研究

https://doi.org/10.1002/imt2.21

5.4
iMeta
RESEARCH ARTICLE
● 2022年5月4日,华南农业大学兽医学院曾振灵团队在iMeta在线发表题为“The co-occurrence of antibiotic resistance genes between dogs and their owners in families”的研究性文章。
● 该研究主要对宠物犬与宠物犬主人的亲密关系对他们的肠道菌群和耐药基因进行分析,研究家庭中宠物犬与其主人在抗生素耐药性方面的相关性。指出伴侣动物存在抗生素耐药性并威胁公共健康的风险,因此应警惕在伴侣动物中抗生素的使用。
● 第一作者:赵若男(华南农业大学兽医学院)
● 通讯作者:曾振灵([email protected]);熊文广([email protected])
● 合作作者:郝杰、杨锦涛、童翠红、谢龙飞、肖丹瑜
摘 要
人类与伴侣动物之间的亲密关系与人类的抗生素抗性(耐药性)有着独特且极其重要的关系。但是,目前仍缺少对伴侣动物与宠物犬主人耐药性的全面分析。我们选择13条宠物犬和16名宠物犬主人以及22条狗场的狗,通过宏基因组来分析宠物犬与宠物犬主人的亲密关系对他们的肠道菌群、耐药基因(ARGs)和可移动遗传元件(MGEs)的影响,并研究家庭中宠物犬与其主人在耐药性方面的相关性。狗的肠道微生物具有更高的耐药水平而宠物犬主人的肠道微生物具有更高的物种多样性。在肠道菌群、耐药基因和可移动遗传元件的组成方面,宠物犬与宠物犬主人比起其它组拥有更高的相似性。从家庭的角度来看,宠物犬与其主人在大环内酯类耐药性基因上有着很强的相关性。我们的研究从群落水平上证实了宠物犬与其主人在耐药基因方面的相关性。这些发现意味着伴侣动物存在抗生素耐药性并威胁公共健康的风险,因此应警惕在伴侣动物中抗生素的使用。
关键词:宏基因组学;伴侣动物;狗;粪便耐药组;肠道微生物组
亮 点
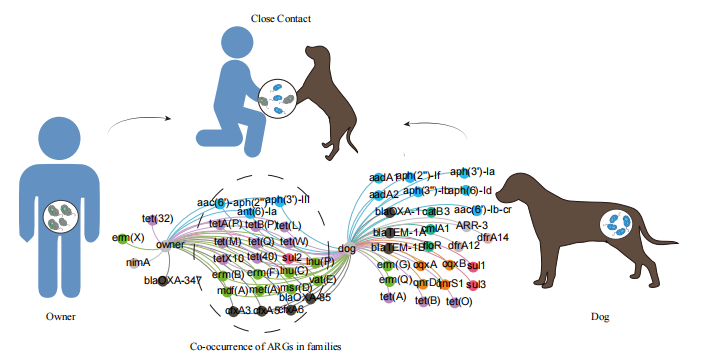
● 宠物犬中较高的变形菌纲导致其更高的耐药水平
● 宠物犬与其主人的亲密关系影响他们的肠道菌群
● 宠物犬与其主人共享的菌群致使家庭中宠物犬和主人的耐药基因共存
视频解读
Bilibili:https://www.bilibili.com/video/BV1aF411L78k/
Youtube:https://youtu.be/1UXxYFPTj2o
中文翻译、PPT、中/英文视频解读等扩展资料下载
请访问期刊官网:http://www.imeta.science/
全文解读
引 言
抗生素耐药性(AMR)被世界卫生组织和疾病控制与预防中心等众多知名组织视为“全球公共问题”。抗生素耐药性和耐药性致病菌引起的感染对人类、动物和环境健康造成了日渐严重的全球威胁。引起感染的病原菌可以在水平基因转移(HGT)的过程中获得耐药基因(ARGs)。由可移动遗传元件(MGEs)介导的HGT是ARGs传播的重要驱动因素,可以促进ARGs传播给受体宿主,从而产生新的耐药菌。MGEs,如质粒,整合性接合元件(ICE),转座子和整合子,可以在DNA分子内或分子间移动,并在细菌间发生转移。
在现代社会中,越来越多的伴侣动物与人类亲密相处。虽然全球各地的伴侣动物的数据有所不同,但是伴侣动物都被主人认为是不可分割的家庭成员。主人可以养伴侣动物甚至只是与它们互动,就可以获得幸福感。此外,在过去的十年里,患有癌症的伴侣动物在人类癌症的药物研发和优化中发挥了重要作用。然而,伴侣动物被认为抗生素耐药性的潜在宿主。众所周知,主人通常与他们的可能携带耐药微生物的伴侣动物处在相同的环境,特别是从One-Health的角度来看,伴侣动物可能是将耐药基因和/或耐药菌传递给主人的源头。来自伴侣动物的主要耐药微生物,包括产碳青霉烯酶的肠杆菌、产超广谱β-内酰胺酶的(ESBL)革兰氏阴性菌、耐甲氧西林金黄色葡萄球菌(MRSA)和耐万古霉素肠球菌,可能直接或间接地对人类健康造成不良影响。据报道,MRSAs可能可以在宠物和主人之间传播。产超广谱β-内酰胺酶肠杆菌的传播对全球人类和兽医医疗环境构成挑战,并威胁公共卫生。欧洲药品管理局(EMA)表明目前对伴侣动物与其主人之间耐药性传播的影响因素和途径缺乏了解。
肠道具有许多抗生素耐药性的决定因素,被视为“肠道耐药组”。目前,只有少数研究表征了伴侣动物的肠道微生物。通过宏基因组分析发现,与其他哺乳动物相比,狗的肠道微生物在基因含量和对饮食的反应方面与人类相似。先前对伴侣动物与其主人之间的耐药性的研究主要依赖培养细菌的方法。然而,基于细菌培养的研究来全面表征伴侣动物和人类的肠道耐药性仍有不足。Belas等人通过基因组方法发现,伴侣动物的耐药组和与其亲密接触的人相似。但是,该研究仅关注与临床重要细菌相关的ARGs,并且由于序列与引物和探针不互补,PCR检测ARGs的能力有限。最近,有研究通过宏基因组组装和分箱方法对抗生素耐药性与微生物群落之间的联系提供新见解。我们在此通过宏基因组组装和分箱方法,来研究宠物犬与主人之间的亲密接触对他们的肠道耐药组的作用,并解读他们之间ARGs的流行性和潜在转移。研究路线如图1所示。
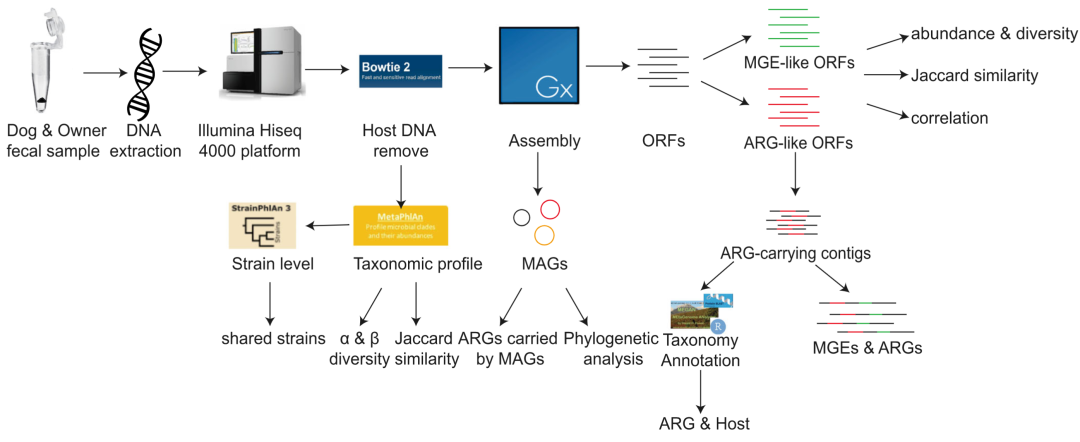
图1. 本研究的分析路线
对16名宠物犬主人、13 条宠物犬和 22 条狗场的狗的粪便样本进行宏基因组测序,结合网上下载的22组Hadza的宏基因组数据,在去除宿主DNA后,进行数据组装和微生物群落的分析。比对resfinder和自制的MGE数据库来检测ARGs和MGEs。比较分析不同组间ARGs和MGEs的总丰度、多样性、Jaccard相似性和相关性。此外,我们解读了ARG的细菌宿主,并注释了周围的MGEs。ARG,抗生素耐药基因; MAG,宏基因组重组装的基因组; MGE,可移动遗传元件; ORF,开放式阅读架
方法
样品采集
从15个家庭分别收集了13条宠物犬和16名宠物犬主人的粪便样本,其中从10号家庭的主人中采集了两份样本。为了验证宠物犬与主人之间的亲密接触对他们耐药组和微生物组的影响,我们从3个不同的狗场中收集了22条很少与人接触的狗的粪便样本,并从网上下载了22组Hadza,一类很少接触抗生素和狗的人群,的宏基因组序列数据。表S1总结了这些样品的详细信息。我们将采集到的所有粪便样本保存在4 ℃,并在2天内运输到实验室,然后在提取DNA前储存在-80 ℃。
DNA的提取
每个样品取0.25g,通过MoBio PowerSoil DNA分离试剂盒提供的方法提取DNA。DNA的纯度和浓度由NanoDrop ND-2000仪器(NanoDrop Technologies)通过紫外光谱法测定。将OD260/OD280在1.8 – 2.0范围和浓度>50 ng/μl的DNA样品用于宏基因组测序
宏基因组测序和质检
在测序前,构建大小约为300bp 的DNA片段文库。每个粪便样本使用Illumina HiSeq 4000平台进行 2×150 PE 的双端测序。使用Sickle去除平均质量Q < 20或长度L< 20 bp的原始数据。
宿主DNA的去除、宏基因组装和分箱
用参考基因组CanFam3.1(NCBI号GCA_000002285.3)和GRCh38.p13(NCBI号NC_000001.11)通过bowtie2-build分别创建狗和人的参考基因组数据库。使用创建的参考基因组数据库,处理质控后的数据来去除宿主DNA。表S1总结了每个宏基因组样本的原始数据(raw reads)和净数据(clean reads)大小。使用 CLC Genomics Workbench(v 10.01)软件根据默认参数 k-mer值对clean reads从头开始组装的。通过metaWRAP(v 1.2.1)软件中的分支算法“MetaBAT2”模块对重叠群contigs进行深度组装来获得宏基因重组的基因组(MAGs)。使用CheckM v1.1.2评估获得的MAGss的完整度和污染度[54]。仅保留完整度≥50%,污染度<10%的MAGs
ARG和MGE检测
使用Prodigal(v 2.6.3)对组装后的contigs进行开放阅读框(ORF)的预测。将预测得到的ORFs的核苷酸序列分别与Resfinder数据库和自建的MGE数据库进行BLASTN比对,将相似度≥80%和序列覆盖度≥70%的ORF序列视为类耐药基因开放阅读框(ARG-like-ORFs),将相似度≥90%和至少25个氨基酸匹配上的ORF序列视为类可移动遗传元件开放阅读框(MGE-like-ORFs)。使用CLC软件,通过将ARG-like-ORFs和MGE-like-ORFs分别映射到相应的clean reads,根据参数:最小长度覆盖长度≥ 95%;相似度≥95%,计算具有95%相似性的最小长度覆盖率为95%的重叠群和相邻群最小长度≥500 bp的重叠群来计算ARG-like-ORFs和MGE-like-ORFs的覆盖度。每个宏基因组样本中的细胞数通过ARGs-OAP(v2.0)中计算。每个样品的ARG-like-ORFs和MGE-like-ORFs的丰度根据以下公式进行标准化处理。
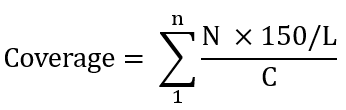
这里,N属于映射到ARG-like-ORFs或MGE-like-ORFs的次数,L是目标ARG-like-ORFs或MGE-like-ORFs序列长度,n是ARG-like-ORFs或MGE-like-ORFs的数量,150是Illumina测序的读长,C表示每个样品的细胞数。
MAGs携带的ARGs的鉴定同样遵循上述方式。为了进一步评估ARGs的转移,通过ICEfinder在线工具,使用默认参数预测了携带ARGs的MAGs中的ICEs和IMEs。那些携带整合酶、松弛酶和T4SS基因簇的元件被定义是T4SS型ICEs,而没有T4SS但具有整合酶和松弛酶的元件被确定为IMEs
肠道菌群分析
使用MetaPhlAn 2对去除宿主DNA后的clean reads进行菌群结构注释。使用StrainPhlAn3进行菌株水平分析和菌株跟踪分析。我们利用StrainPhlAn3获得了来自伴侣动物的主要耐药性微生物的共识菌株。除此之外,我们将含有ARGs的contigs预测ORFs,用其蛋白序列与NCBI 无冗余蛋白库(NRDB)进行BLASTP比对,将BLASTP的结果使用MEGAN(v5,MEtaGenome ANalyzer)根据默认参数进行注释。,如果一个contig中有一半以上的ORFs来自一个物种,便将其注释为该物种,否则,它被视为未分类。
MAG的系统发育分析和分类分析
使用基因组分类数据库工具包(GTDB-Tk)对筛选留下的MAGs进行注释,使用PhyloPhlAn 3.0软件进行系统发育树分析。
统计学分析
在多组比较中使用非参数Kruskal-Wallis检验,在家庭中宠物犬与其主人的比较用非参数Mann-Whitney U检验,通过Bonferroni完成多次测试校正的。P < 0.05被认为具有统计学意义。ARGs、MGEs和物种组成的香农多样性在R(v4.0.2)计算完成。箱线图在Origin(v9.6.5)完成。网络图的可视化在Cytoscape(v3.7.1)和Gephi(v0.9.2)平台上完成。使用iTOL v5在线绘制系统发育树。线性回归在网站:https://hiplot.com.cn完成。剩下的的都是在R或ImageGP网站完成的。
结果
狗的肠道具有高水平的ARGs
我们比较了宠物犬、狗场的狗、宠物犬主人以及Hadza四个组的ARG,MGE和物种组成来探究他们的差异性。宠物犬、狗场的狗的肠道微生物比宠物犬主人和Hadza的肠道微生物具有更高的ARGs丰度。但是,宠物犬、狗场的狗和宠物犬主人的MGEs的丰度没有显著差异(图2A,B)。在ARGs和MGEs的α多样性方面的结果跟上述一致。但是,宠物犬和狗场的狗的物种多样性明显低于宠物犬主人(图2C,D和S1A)。Hadza不仅在ARG和MGE的丰度上是最低,而且在ARG,MGEs和物种多样性上也是最低的。
对宠物犬、狗场的狗、宠物犬主人以及Hadza四个组的肠道微生物,ARGs和MGEs通过PCoA进行聚类分析。我们发现宠物犬和宠物犬主人不仅在ARG还在MGE组成上聚类成一组(P > 0.05,Adonis),但是在它们的物种组成上,他们具有明显的差异(P < 0.05,Adonis)(图2E,F和S1B),这种情况也出现在宠物犬和狗场的狗的中。狗场的犬和宠物犬主人的不仅在ARG还在物种组成上存在明显差异(P < 0.05,Adonis)。Hadza在物种、ARG和MGE组成上与宠物犬,宠物犬主人和狗场的狗都所不同(P < 0.05,Adonis)。
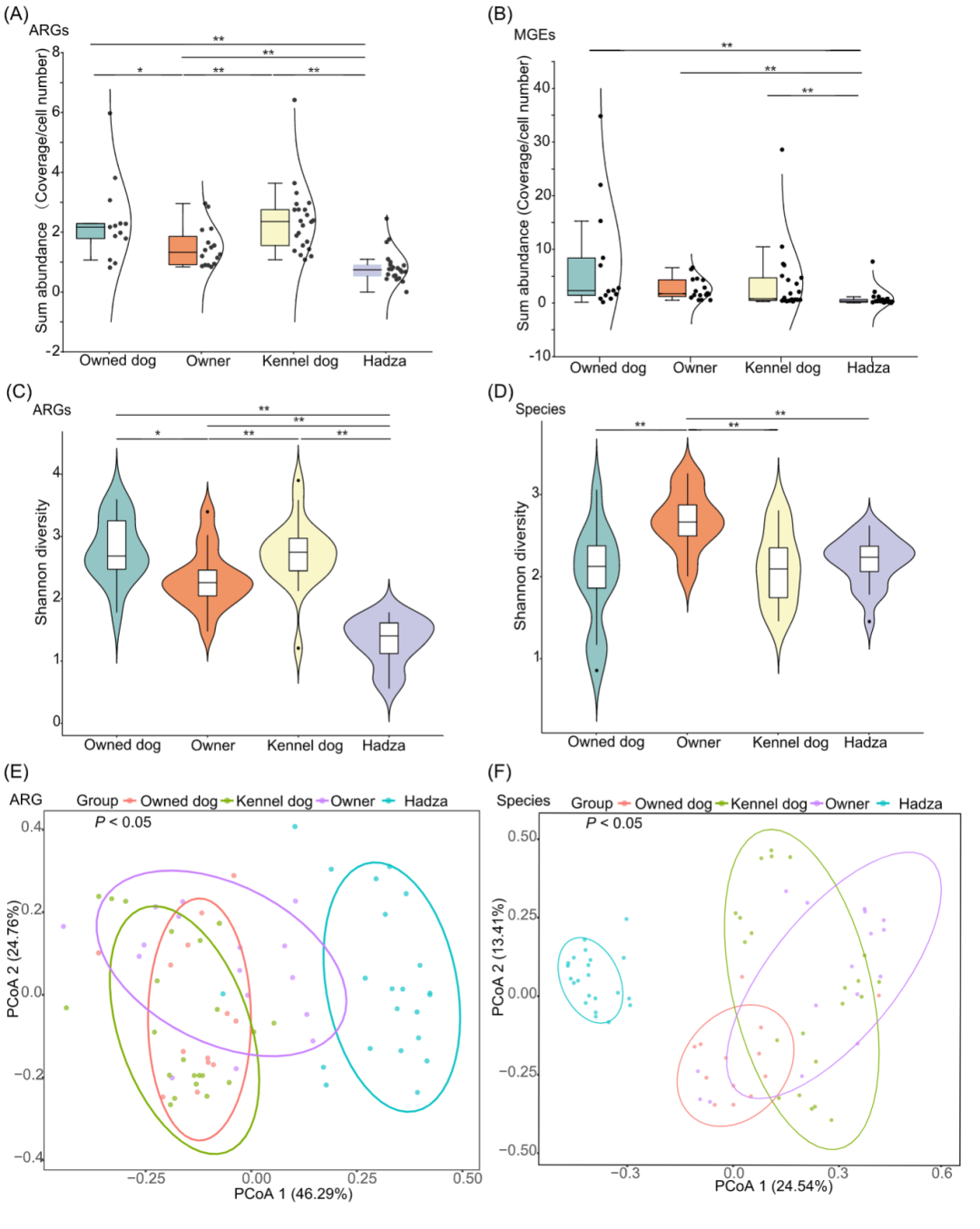
图2. 宠物犬、狗场的狗、宠物犬主人以及Hadza的ARGs和MGEs的总丰度和多样性
(A)ARGs总丰度;(B)ARGs总丰度;(C)ARGs的α多样性;(D)物种的α多样性。(E)肠道ARGs的 PCoA。(F)肠道微生物的PCoA。宠物犬(n = 13),宠物犬主人(n = 16),狗场的狗(n = 22),Hadza(n = 22)。ARG,抗生素耐药基因; MGE,可移动遗传元件; PCoA,主坐标分析。*p < 0.05, **p < 0.01
宠物犬与主人具有相似的微生物和耐药基因组成
我们分别统计了宠物犬、狗场的狗、宠物犬主人以及Hadza四组共享的ARG,MGEs和菌群数量,并使用Jaccard指数进一步分析了他们在物种、ARG和MGE组成上的相似性。宠物犬与主人共享了70%的ARG和82%的MGE,共享的ARG和MGE数量在宠物犬主人中分别占比为72%和82%。狗场的狗拥有最多种类的ARGs和MGEs。但是,狗场的狗仅与宠物犬共享57%的ARGa,且狗场的狗和宠物犬主人之间共享的ARGs仅占其总ARGs的52%。狗场的狗分别与宠物犬和宠物犬主人共享的MGE的比例也低于他们所共享的MGEs的比例(图3A,B)。此外,在宠物犬的肠道中发现的81%的物种在宠物犬主人的肠道中同样也检测到(图3C)。宠物犬和主人之间的ARGs,MGEs和肠道菌群的Jaccard指数分别为0.55,0.69和0.55。与其他组相比,宠物犬和主人之间的肠道物种、ARG和MGE组成上的相似性更高(图3D-F)。
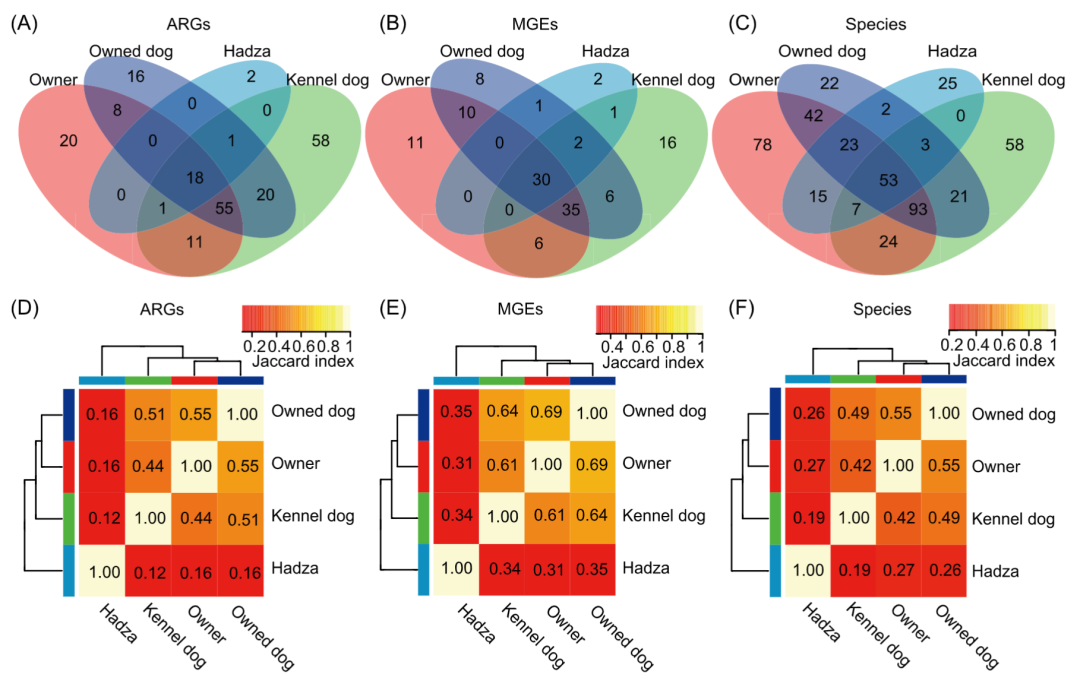
图3. 宠物犬、狗场的狗、宠物犬主人以及Hadza人在肠道ARG、MGE和物种方面的关系
(A–C)这四组中共享的ARGs(A),MGEs(B)和物种(C);(D–F)肠道ARGs(D),MGEs(E)和物种(F)的距离矩阵热图。物种由 MetaPhlAn 2 定量。ARG,抗生素耐药基因; MGE,可移动遗传元件
家庭中宠物犬与主人的比较
接着,我们从家庭的角度挑选了12对宠物犬和主人,来比较他们的肠道物种、ARG和MGE的组成。同样地,我们发现宠物犬的ARG总丰度和香农多样性均高于主人,而物种香农多样性要低于主人;两组之间的MGEs总丰度和香农多样性没有显着差异。但是,我们发现在β多样性方面观察到与先前不一样的结果。宠物犬和主人不仅在物种组成方面,还在ARG组成方面存在差异(P < 0.05,Adonis;图4A,B)虽然,在MGE组成上,他们之间没有显着差异(P > 0.05,Adonis;图 S2A)。宠物犬和主人在ARG和MGE组成的差异同样也体现在在ARG和MGE大类上(图S2B,C)。在ARG大类上,宠物犬和主人的前四种ARG大类是氨基糖苷类,β-内酰胺类,大环内酯类和四环素类。这四种ARG大类的丰度在宠物犬和主人的ARG总丰度中的占比分别为83%和95%。宠物犬和主人中丰度最高的ARG大类都是大环内酯类,其丰度在宠物犬狗中是高于主人的。此外,氨基糖苷类和甲氧苄啶类耐药基因的丰度在宠物犬中也高于主人。当在MGE大类方面,转座酶在所有样品中是最常见的MGE类型。与主人相比,宠物犬的整合子相关整合酶和qacEdelta的丰度更高(图4C)。在纲水平上,γ-变形菌纲和ε-变形菌纲在狗中更常见,而主人有更多的梭菌纲和δ-变形菌纲(图S2D)。在科水平上,拟杆菌科在粪便样本中丰度最高,其丰度在主人中高于宠物犬。宠物犬的肠杆菌科和消化链球菌科的丰度更高(图4D)。此外,我们通过使用线性判别分析(LEfSe)表征了宠物犬和主人的粪便微生物群的差异,并发现了指示属(P < 0.05)(图S3)。
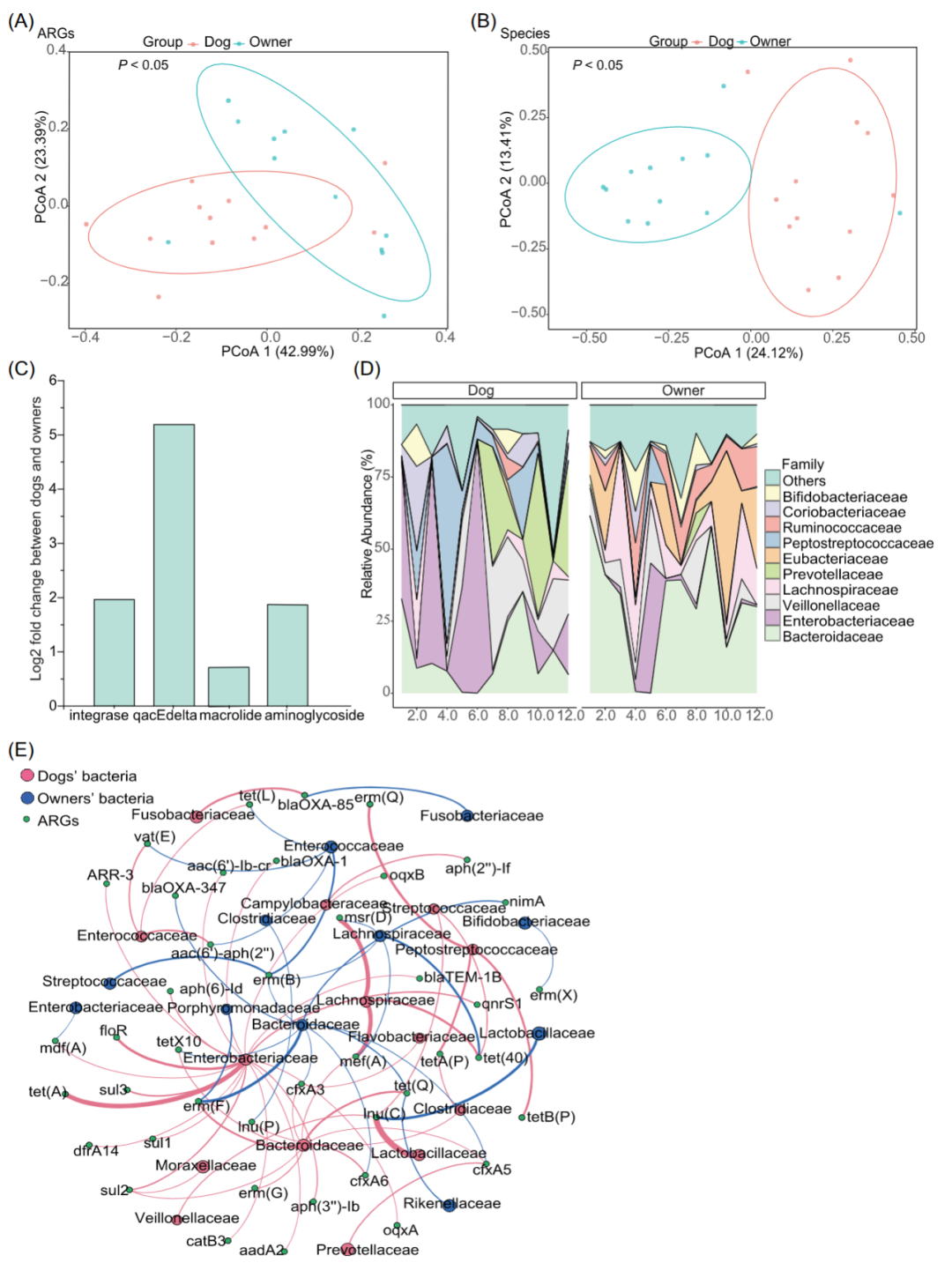
图4. 宠物犬和主人的ARG,MGE和物种组成上的比较
(A)肠道ARGs的 PCoA;(B)肠道微生物组的PCoA;(C)在ARG和MGE大类上的差异;(D)前10的科。其余表示剩下的科;(E)ARGs及其在科水平的细菌宿主。仅显示相对丰度超过0.01的ARGs。节点根据模块化类进行着色。曲线的大小代表了细菌宿主携带的ARGs丰度。宠物犬(n = 12) 和主人(n = 12)。ARG,抗生素耐药基因; MGE,可移动遗传元件; PCoA,主坐标分析
肠道耐药组与微生物组的组成相关
我们通过普鲁克分析(Procrustes)探索了肠道ARG、MGE和微生物的组成之间的联系,分析结果表明,ARG、MGE和微生物的组成彼此显着相关,表明肠道微生物构建了ARGs和MGEs(图S4)。我们通过计算Pearson相关系数,检测并总结了哪些微生物(在不少于五个样本中)与ARGs和MGEs的总丰度相关(表S2)。我们发现在宠物犬和主人中都观察到了大肠杆菌,埃希菌属,肠杆菌科和变形菌等与ARGs和MGEs的总丰度之间存在很强的相关性。
同时,我们通过对携带ARGs的contigs进行分类注释来解读ARGs的细菌宿主。在科水平上,肠杆菌科是宠物犬主要的细菌宿主,并拥有最种类的ARGs,包括宠物犬的大多数ARG大类,而拟杆菌科是主人的ARGs的主要细菌宿主(图4E)。在纲水平上,变形菌纲携带了宠物犬的大部分不同种类的ARGs,而主人的ARGs主要由拟杆菌纲携带。
家庭中ARGs和菌株的共存
我们分析了宠物犬和主人在ARGs和共享菌株上的相关性,并总结了位于同一contig上的ARG周围的MGEs,以探索ARGs在家庭中传播的可能性。在宠物犬和主人中分别共检测到118和89种ARGs。其中超过一半的ARGs(61个)由宠物犬和主人共享。这些共享的ARGs在家庭环境中普遍存在,在宠物犬和主人的ARG总丰度中分别占比88%和96%。我们还发现,共享ARG的丰度与ARGs总丰度显着正相关(主人r = 0.988,狗r = 0.925,p<0.01)。采用二分图展示宠物犬与主人共享的ARGs以及他们拥有的独特ARGs(图5A)。大部分共享的ARGs属于常用抗生素的耐药基因,如β-内酰胺,氨基糖苷类和四环素,大环内酯类。在共享的ARG中, tet(Q)、 tet(A)、 lnu(C)、mef(A)、erm(F)和erm(B) (对四环素和大环内酯类耐药)等ARGs的丰度在宠物犬和主人中占比最大。使用冗余分析(RDA)研究了共享的菌群(在纲水平上),MGEs和ARGs之间的关系(图S5B)。为了确定共享的菌群和MGEs对共享的ARGs的贡献,进行了方差分解分析(VPA),我们发现所选择的因子可以解释87.52%的共享的ARGs。共享的菌群和MGEs的共同作用对共享的ARGs的贡献度为28.36%,共享的菌群对共享的ARGs单独贡献了45.40%,高于共享的MGEs的单独作用(13.76%)(图5B)。我们还利用StrainPhlAn3来研究宠物犬及主人是否存在共享的菌株,在所有的粪便样本中发现了12个大肠杆菌的共识菌株,其中有两对来自同一家庭(图5C)。此外,还获得了四个肺炎克雷伯菌的共识菌株(图S6A)。此外,我们计算了宠物犬和主人之间ARGs的Pearson相关系数。虽然在ARGs的总丰度上,宠物犬和主人没有显示出明显的相关性(P > 0.05),但我们发现他们的大环内酯类耐药基因(r = 0.92,P < 0.01)表现很强的相关性,在ARGs上也观察到了相关性,如mdf(A)和tet(L)(图5D和S6B)。
ARGs与MGEs组成也显着相关(图S4)。我们调查了来自同一家庭的宠物犬和主人共享的ARG的周围MGEs。从样品中共发现了104个携带至少一个ARG和MGE的contig(表S3),在这104个contigs中,有6个具有相同基因排列的contig由来自同一家庭的宠物犬和主人共享,35个contigs由宠物犬和主人共享,最常见的contigs是同时携带cfxA3和tnpA。除此之外,我们发现好几个携带ARGs的contigs物种归类到大肠杆菌
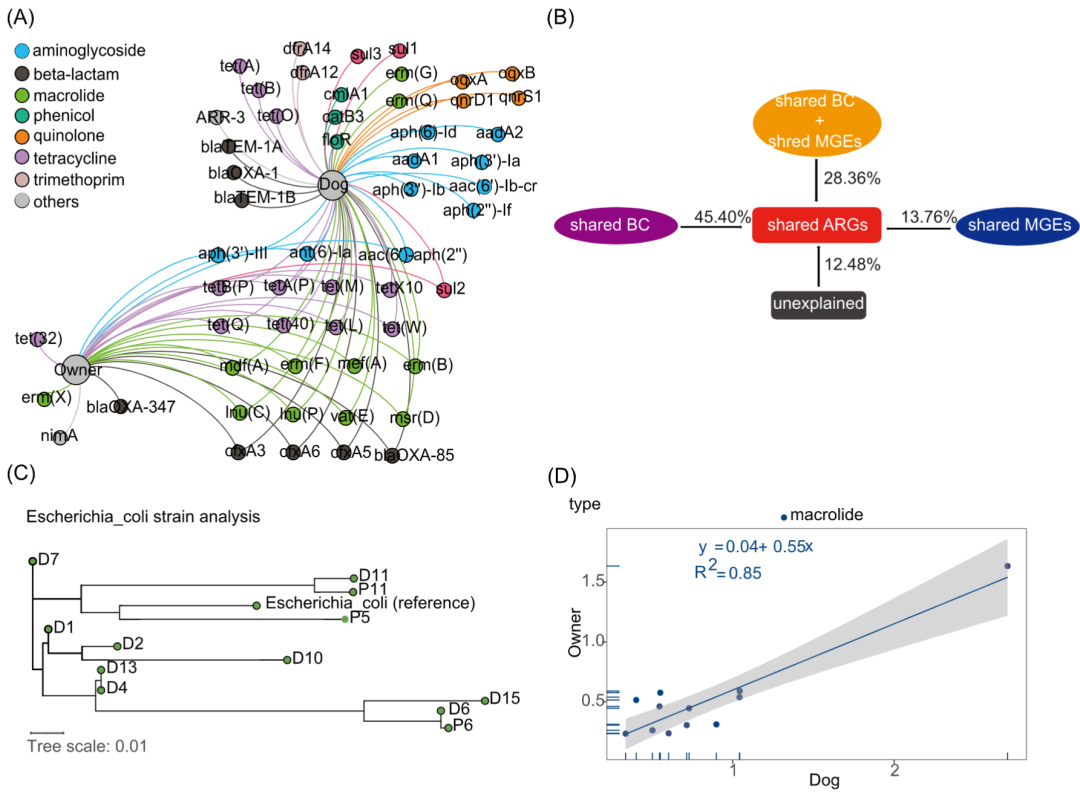
图5. 对宠物犬与主人之间共享的ARGs和菌株的分析
(A)宠物犬和主人共享ARG的二分图。仅显示相对丰度超过0.01的ARGs。节点根据模块化等级着色,曲线的大小表示具有该ARG的样本数量;(B)方差分解分析展示了共享的菌群和MGEs对共享的ARGs的贡献度;(C)使用StrainPhlAn3分析宠物犬和主人的大肠杆菌系统发育树。红色方框表示来自同一家庭的宠物犬和主人。大肠杆菌的参考基因组选择的大Escherichia coli str. K-12 substr. MG1655;(D)宠物犬和主人在ARG大类上的相关性。ARG,抗生素耐药基因; MGE,可移动遗传元件
MAGs揭示了具有代表性的菌及其携带的ARGs
使用分箱得到重组的基因组来分析代表性细菌及其携带的ARGs。从来自12个家族的24个样本中,通过重组获得细菌基因组,分别在宠物犬和主人中获得215和326个MAGs。我们将宠物犬和主人的MAG组合在一起进行系统发育树分析。系统发育树显示它们属于六个菌门,包括放线菌门,拟杆菌门,广古菌门,厚壁菌门,梭杆菌门和变形菌门(图6A)。由70个MAGs被鉴定到种在物种水平上分类,58个MAGs被鉴定到属,224个MAGs被鉴定到科,124个MAGs被鉴定到目,6个MAGs被鉴定到纲,29个MAGs被鉴定到门,剩下的20个MAGs由于可用相关参考基因组的限制仅被鉴定到细菌。
在获得的541个MAGs中,共有58个MAG(宠物犬30个,主人28个)被确定为携带ARGs(n = 108)。携带ARGs的细菌宿主在宠物犬和主人中主要归属于肠杆菌科(图6B)。58个MAGs共携带9种类型的ARGs,最常检测到的4种ARG大类分别是大环内酯类,四环素,β-内酰胺和氨基糖苷类。此外,由21个MAGs(宠物犬11个中,主人10个)携带至少两个ARGs。在主人中注释为肠杆菌科的两个MAGs携带了五个ARGs,它们分别对β-内酰胺(blaSHV-187和blaSHV-199),磷霉素(fosA和fosA6),大环内酯(mdf(A))和喹诺酮(oqxA和oqxB)类抗生素耐药。此外,注释到拟杆菌科的MAGs总共由13个(宠物犬4个,主人9个),它们携带的ARGs对大多数抗生素类型包括氨基糖苷类,β-内酰胺,大环内酯类,磺胺类和四环素等耐药。
我们进一步预测了携带ARGs的MAG中s的ICE以及综合和可移动元件(IME),以进一步估计ARG的可传播性(表S4)。在22个携带ARGs的MAGs中共鉴定出13个T4SS型ICE和30个IME,其中6个MAP同时包含ICE和IME区域。对于其他组分,发现五个MAGs中的七个IME含有oriT区域,这是一个短的DNA序列,对接合至关重要。此外,还观察到大多数ICE和IME区域两侧的附着位点(attL和attR),表明它们在特定位点重组的潜在作用.
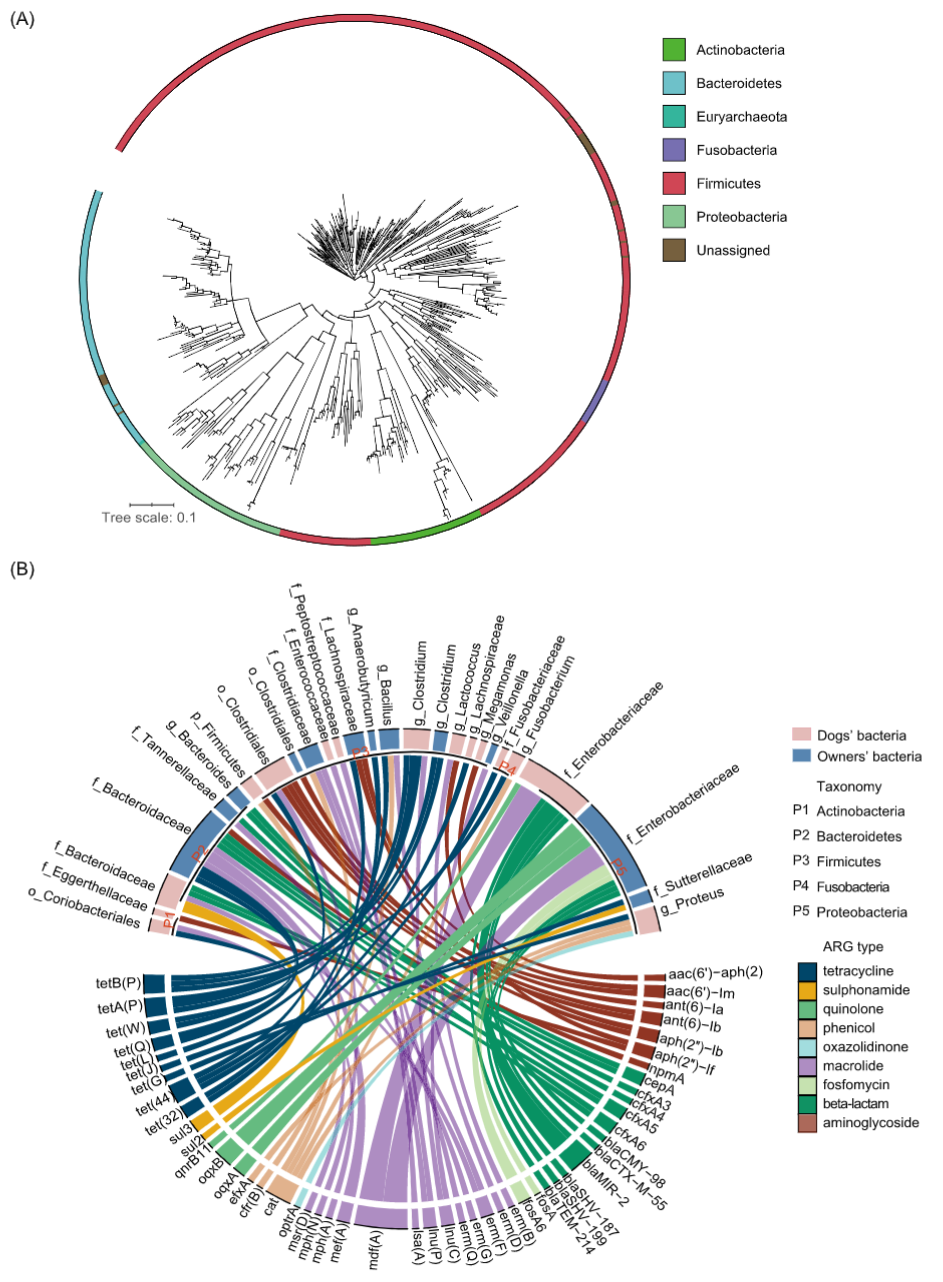
图6. MAGs分析
(A)MAGs的系统发育树,根据门的类别着色;(B)携带ARGs的MAGs与携带的ARGs的映射,内环长度分别代表ARGs和MAGs的数量
讨论
伴侣动物与其主人的亲密接触具有传播耐药性微生物的风险。特别是位于宠物犬皮肤和黏膜上的耐药性微生物很容易转移到主人身上。此外,一些耐药情况令人担忧,因为有对粘菌素耐药的mcr-1可能从伴侣动物传播给人类事件的报道[23]。
在这项研究中,我们报道了宠物犬、主人、狗场的狗和Hadza在肠道ARG,MGE和菌群组成上的比较。目前的研究主要使用传统的细菌培养方法来探索狗和人类的耐药性微生物。然而,这些方法的局限性在限制了在社区范围内的研究。使用宏基因组组装和分箱的方法,我们全面了解了宠物犬和主人之间耐药性微生物的相关性。狗的肠道具有更高的耐药水平,而宠物犬主人具有更丰富的物种组成,但是他们在MGEs方面没有显着差异。宠物犬和狗场的狗有更高的耐药水平,可能是由于抗生素的滥用。抗生素是治疗宠物传染性细菌性疾病的有用工具,然而,对于宠物抗生素的规范使用尚无明确指南。小动物的抗生素的使用已被确定为耐药性微生物定植或感染的危险因素之一。此外,耐药菌和ARGs还可以来源它们的食物。对于狗场的狗,另一个可能性原因是圈养。
最近的研究表明,狗从野生和未驯化到与人类同居的生活方式的转变已经改变了家养的狗的肠道菌群。与其他哺乳动物相比,人类和狗的肠道微生物之间存在许多相似之处。我们发现,宠物犬不仅在肠道微生物方面与宠物犬主人更相似,而且在ARG和MGE方面都与宠物犬主人更相似。先前的研究强调了环境在肠道微生物的重要作用,因为他们发现有关联的人类比没有关联的人类在肠道微生物组成上更相似。同样,具有遗传相关的狗比不相关的狗的在肠道微生物组成上更相似。此外,生活在家庭中的狗与生活在庇护所中的狗具有不同的肠道微生物组成,这表明生活环境可能对肠道微生物的组成具有重要影响。同时,宠物和主人之间的亲密关系会影响主人的肠道菌群。与没有养宠物的人类相比,在家庭中养狗通常会导致主人皮肤上的微生物种类更丰富。我们的研究结果加强了宠物犬与主人同居生活以及他们的亲密关系在肠道微生物群调节中起着重要作用,并对双方的健康产生影响的观点。
考虑到在人类和伴侣动物的亲密关系以及检测到耐药菌的频率的增加,为耐药性微生物在种间传播创造了新的机会。我们研究了家庭中宠物犬与主人之间耐药性微生物的联系。从家庭的角度来看,宠物犬和它们的主人在ARG和MGE方面有相似之处。大环内酯类耐药基因是宠物犬和主人中丰度最高的ARG大类,而大环内酯类抗生素是人类所用的最多的三大抗生素类别之一。在宠物犬和主人中共检测到18个四环素抗性基因,包括存在的最广泛和最主要,也是肠道微生物携带的典型ARG,tetX。它们在宠物犬中的流行可能是由于伴侣动物的从业者更喜欢使用人类抗生素,因为它们的质量更好,更容易获得。在伴侣动物中,广谱抗菌素和重要的人类抗生素的使用率也很高。
肠道微生物是ARGs的源头,也是耐药率越来越高的致病菌的潜在储存库。ARGs可以通过特定细菌在不同环境之间传播。我们通过Procrustes分析来探探究在细菌种群中潜在的水平基因转移,并发现ARG,MGE和肠道菌群彼此显着相关。以前的研究主要通过网络分析来探索ARGs的潜在宿主。然而,这些基于数学统计的结果仅揭示了有限的ARGs的宿主,且需要进一步验证。最近,几项研究通过注释携带ARG的contigs的ORFs。在此研究中,我们不仅通过相关性分析找出了哪些微生物群与ARG和MGE总丰度显着相关,还通过注释携带ARG的contigs来解读ARGs的潜在宿主。我们发现变形菌纲和肠杆菌科可导致高耐药水平,并且很可能携带大部分不同类型的ARGs。这可以由变形菌纲和肠杆菌科是重要耐药机制传播的关键驱动因素的理论来支持。宠物犬比主人有更多丰度的变形菌纲和肠杆菌科,这表明宠物犬肠道中的基因传播的可能性更高,因为更高丰度的变形菌纲和肠杆菌科可以促进ARGs的传播。结合这一状况,在家庭中,宠物犬作为耐药菌的储存库这一角色引人担忧。
家庭中,宠物犬和主人共享的ARGs在总的ARGs占比很大,并且其高流行率令人担忧。我们的研究结果表明,在家庭中,共享的菌群在宠物犬和主人ARGs的共存中起着重要作用。利用StrainPhlAn3,我们发现大肠杆菌是宠物犬和主人共识菌株。在ARGs方面,我们发现大环内酯类耐药基因和一些特定的ARGs在宠物犬和主人之间显示出很强的相关性。人类和伴侣动物使用几种抗菌药物类型相同可导致检测到的ARGs重叠。对携带ARG和MGE的contig的调查发现,宠物犬和主人有相同的基因排列,这表明这些ARGs通常与MGEs相关,而这对于宠物犬和主人之间的耐药性微生物的传播很重要。我们的研究结果还表明,ARGs和MGE有可能在同一家庭中的宠物犬和主人之间传播。携带ARG的contigs大部分归属到大肠杆菌,表明宠物犬和主人肠道中菌株携带的许多ARG是可传播的。这与上述的大肠杆菌与ARG和MGE总丰度具有很强的相关性的结果一致。但是,由于低丰度的短序列组装的技术限制,基于基因注释的分析无法区分遗传元件的核苷酸水平变异。
宏基因组分箱有助于更好地了解耐药性微生物,并为ARGs的传播提供重要见解。我们通过筛选过的MAGs探究了ARGs的细菌宿主和可移动性。肠杆菌科携带大部分ARGs,这与上述对ARGs细菌宿主的鉴定一致。关于ARGs的可移动性,ICEs作为不同细菌物种和属之间耐药性微生物传播的载体发挥了重要作用被广泛报道。除了ICE外,在一些携带ARGs的MAGs中还发现了染色体源的IME。在一项综述中,IME被认为是ARG的可能储存库。携带多种类型的ARG和ICE地MAGs由于它们会增加ARGs的可移动性和传播风险,应该引起极大的关注,因为。需要进一步的研究来确定宠物犬和主人之间抗生素耐药性传播的因果关系和方向。为了防止抗生素耐药性在家庭中蔓延,制定有效措施来控制抗生素耐药性,需要社会各界的共同努力。
结论
我们首次使用宏基因组组装和分箱方法来探索宠物犬和主人之间的肠道菌群和耐药基因的流行情况和联系。宠物犬拥有更高丰度的变形菌纲和肠杆菌科的丰度较高可能是导致宠物犬高水平耐药的原因。宠物犬和主人之间的亲密关系在他们的肠道耐药组和微生物组的调节中发挥了关键作用,并对他们的的健康产生影响。从家庭的角度来看,共享的肠道菌群可能是家庭中宠物犬和主人耐药基因共存的主要原因
代码和数据可用性:
本研究中使用的数据和脚本可以在以下位置找到:https://github.com/zrn327/close-contact-affect-gut-bacterial-community。本文所用到的宏基因组数据可以在NCBI序列储存库种找到(接入号:PRJNA721002)。所有的补充材料(文本、图、表、中文翻译版本或视频)也可从线上获取。
引文格式:
Zhao, Ruonan, Jie Hao,Jintao Yang, Cuihong Tong, Longfei Xie, DanyuXiao, Zhenling Zeng, and Wenguang Xiong. 2022.“The Co‐Occurrence of Antibiotic ResistanceGenes Between Dogs and their Owners inFamilies.”iMeta,1:e21.https://doi.org/10.1002/imt2.21
作者简介

熊文广(通讯作者)
● 华南农业大学兽医学院副教授
● 依托国家兽医微生物耐药性风险评估实验室和华南农业大学兽药研究评价中心,开展畜禽肠道耐药病原菌的传播机制与控制技术、兽药安全评价方面的研究工作,主持国家自然科学基金、国家重点研发计划子课题等项目,相关学术成果发表于Microbiome、iMeta、Environment International等期刊

曾振灵(通讯作者)
● 华南农业大学兽医学院教授,兼任中国畜牧兽医学会兽医药理毒理学分会名誉理事长等职务
● 长期从事兽医药代动力学、新兽药研发与安全评价等方面的工作。发表SCI论文150余篇,出版专著、教材等10余部,获授权发明专利20多项
更多推荐
(▼ 点击跳转)
iMeta|南医大孔祥清团队创建前瞻性多组学纵向原发高血压队列eHypertension
iMeta | 深圳先进院马迎飞组开发基于神经网络分析肠道菌群的方法
iMeta | 南医大陈连民等综述从基因组功能角度揭示肠菌对复杂疾病的潜在影响
iMeta | 北大陈峰组综述口腔微生物组的标准化研究:从技术驱动到假说驱动
iMeta | 电子科大林昊组开发蛋白质赖氨酸乳酸化位点预测工具DeepKla
iMeta | 南昌大学丁霞等-水产养殖系统对中华鳖微生物组和肠道代谢组的影响
iMeta | 华中科大宁康组综述用于蛋白质结构预测的宏基因组定量分析
iMeta | 中科院李小方等膳食甘草促进小鼠镉解毒并调节肠道菌群代谢
iMeta | 浙大倪艳组MetOrigin实现代谢物溯源和肠道微生物组与代谢组整合分析
iMeta | 南科大宋毅组综述逆境胁迫下植物向微生物组求救的遗传基础(附招聘)
期刊简介

“iMeta” 是由威立、肠菌分会和本领域数百位华人科学家合作出版的开放获取期刊,主编由中科院微生物所刘双江研究员和荷兰格罗宁根大学傅静远教授担任。目的是发表原创研究、方法和综述以促进宏基因组学、微生物组和生物信息学发展。目标是发表前10%(IF > 15)的高影响力论文。期刊特色包括视频投稿、可重复分析、图片打磨、青年编委、前3年免出版费、50万用户的社交媒体宣传等。2022年2月正式创刊发行!
联系我们
iMeta主页:http://www.imeta.science
出版社:https://onlinelibrary.wiley.com/journal/2770596x
投稿:https://mc.manuscriptcentral.com/imeta
邮箱:[email protected]
微信公众号
iMeta
责任编辑
微微