人类基因和基因组
基因及其结构
- 基因是细胞内遗传物质的结构和功能单位
- 以脱氧核糖核酸(DNA)的化学形式存在于染色体上
DNA分子组成
- 基本单位为 脱糖核苷酸
- 脱氧核苷酸的组成为 一分子脱氧核糖 一分子磷酸 一分子含氮碱基
- 碱基有四种 == A腺嘌呤T胸腺嘧啶C胞嘧啶G鸟嘌呤==
- 4种脱氧核苷酸按一定顺序首尾相接构成脱氧多核苷酸单链
- 每条脱氧核苷酸单链都有3’端 (-OH)和 5‘端 (磷酸基团)
- 相邻脱氧核苷酸通过磷酸二酯键相连
DNA分子双螺旋结构
- DNA由两条碱基互补反向平行排列 脱氧多核苷酸单链组成
- 一条5’-3’ 方向 一条 3’-5’方向
- AT之间2个H键相连 CG之间3个H键相连
- 两条互补链围绕一个“主轴”向右盘旋双螺旋结构
- 双螺旋表面形成两条凹槽 分为大沟小沟
- 两条沟 对有特定功能蛋白质或酶识别调节DNA上遗传信息非常重要
- DNA上碱基排列组合方式无限
基因结构
- 绝大多数真核生物编码蛋白质的基因为 割裂基因
- 割裂基因由 编码序列(外显子exon)和非编码序列(内含子intron)组成 两者相间排列
侧翼序列
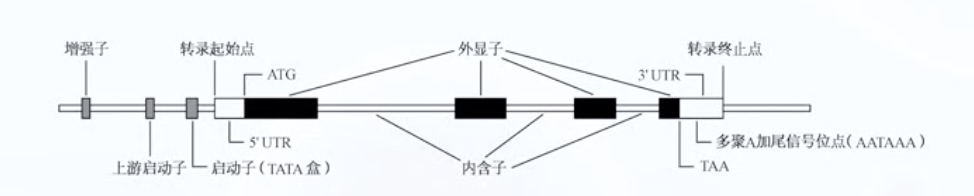
- 每个割裂基因在第一个和最后一个外显子的外侧 都有一段不被转录的非编码区 为侧翼序列
- 侧翼序列包含 启动子 增强子 以及 终止子, 它们对基因的有效表达起调控作用
启动子
- 位于基因起始密码子上游
- 与RNA聚合酶结合而启动基因的转录
- 常见的启动子序列包括TATA框 CAAT框 和GC框
增强子
- 本身不具启动子活性
- 可增强基因转录活性的一段特定的DNA序列
- 位置较自由
终止子
- 位于基因末端的一段回文序列 (反向重复序列 转录可形成发卡式结构)
- 与特定AATAAA顺序组成 阻碍RNA聚合酶移动
- 使酶从DNA模板上脱落 终止转录
外显子和内含子的接头区
- 高度保守的一致序列 成为外显子-内含子接头
- 每个内含子两端具有广泛同源性和互补性
- 5’ 端起始的两个碱基是 GT
- 3’端最后的两个碱基是AG
- 这种接头形式为 GT-AG法
基因组的组成
人类基因组
- 人体所有遗传信息的总和
- 包括相对独立又互相关联的 核基因组和线粒体基因组
- 不特殊说明 通常指核基因组
- 人类基因组约有2w-2.2w个基因
- 与蛋白质合成有关的为1.1%
- 4%为基因调控序列 和 RNA基因序列
- 20% 为内含子、基因非翻译区序列及假基因
- 75%为基因外序列
- 55% 为重复DNA序列
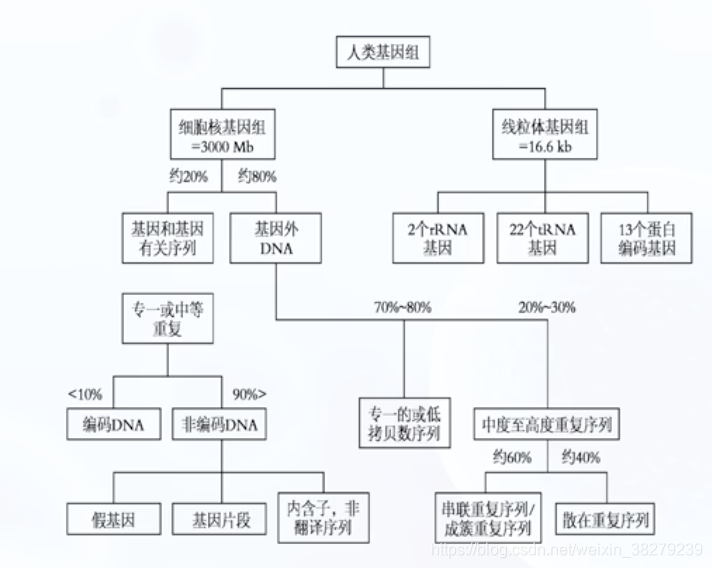
DNA序列的分类
- 根据DNA序列在基因组出现的次数多少
- 可划分为 单拷贝序列 和 重复多拷贝序列
单拷贝序列
- 在基因组中只出现一次或少数几次
- 大多数编码蛋白及酶类基因及结构基因为单一序列 即非重复序列
- 单拷贝或低拷贝序列可占人类基因组大约45%
重复多拷贝序列
- 在基因组中有很多拷贝的DNA序列
- 根据重复序列来源分布特点 可分为 串联重复序列 和 散在重复序列
串联重复序列
- 以5bp 10bp 20bp 或 200bp为一个重复单位 重复很多次 约占整个基因组10%
- 卫星DNA
- 重复次数多 长度达几百kbp
- 异染色质区
- 小卫星DNA 可变数目串联重复 VNTR
- 15bp-100bp的组成单位
- 重复20-50次 形成0.1-20kb的短DNA
- 多数VNTR用于标记
- 少数与疾病相关
- 微卫星DNA 短串联重复 STR
- 基因组的间隔序列 与 非编码区
- 重复序列1-6bp
- 长度小于100bp
- 表现多态性 可拥于基因定位 群体进化 基因诊断的研究
散在重复序列
- 以分散方式分布于整个基因组内的重复序列 约占45%
- 短散在核元件 SINES 100-400bp
- 典型 :Alu 重复
- 人类基因组含量最丰富的散在重复序列
- 平均每3kb发生一次以上重复
- 占基因组总DNA含量11% 长达300bp 重复30w-50w次
- 含有限制性内切酶Alu I的特异性识别位点 AGCT 可被Alu I酶解为170bp和130bp的两个片段
- 长散在核元件 LINES 5k-7kbp
- 如 Kpn I家族
- 可由限制性内切酶Kpn I切割
基因复制与表达
遗传信息的储存单位
- DNA的脱氧核苷酸长链上每3个相邻的碱基序列构成一个三联体 triplet
- 每个三联体密码能编码某种氨基酸
- 也称三联体密码 遗传密码 密码子等
- 4种碱基 4^3 构成64种遗传密码 3个终止子
- 遗传密码在病毒 原核生物 真核生物都是通用的 其中线粒体DNA有3个遗传密码与通用密码不同
- 61种编码氨基酸的密码中,除甲硫氨酸和色氨酸UGG分别仅有一种密码子 其余氨基酸有2-6种密码子
- AUG 若位于mRNA的5’端起始处 即蛋白质合成起始信号 为起始密码子;若不在mRNA起始端 同时还编码 甲酰甲硫氨酸 和 甲硫氨酸
- UAA UAG UGA不编码任何氨基酸 作为肽链合成终止信号
基因的复制
- 复制发生细胞分裂周期 S期
- 以DNA分子自身为模板
DNA复制特点
- 双向复制 以起始点开始 同时向两侧相反方向进行
- 半保留复制 每次复制的DNA中保留一个亲链
- 反向平行 5-3 3-5
- 半不连续复制
- DNA聚合酶只能催化新DNA链沿5-3方向合成
- 由5-3 合成速度快 完成早 为前导链
- 3-5 需先借助RNA聚合酶以DNA为模板 合成长约10bp的RNA 为引物RNA
- 每个引物RNA只合成一个DNA片段 为 冈崎片段
- 此链合成不连续 合成速度慢 为后随链
- 真核生物中冈崎片段100-200bp
- 所有冈崎片段合成后 引物被切除 替换上DNA连接片段 由DNA连接酶将其连接成完整后随链
基因表达
- DNA序列 通过转录 翻译形成具有生物活性蛋白质 从而决定生物各种性状 这一过程为 基因表达
- 转录
- 5-3 链为编码链
- 转录为 以DNA3-5反编码链为模板 按碱基互补配对原则 合成RNA 为hnRNA 不均一核RNA 其经过戴帽 加尾 剪接等加工 才形成mRNA
- 转录最终产物为 mRNA tRNA rRNA
hnRNA加工为成熟的mRNA
- 剪接
转录合成的hnRNA需经过剪接、切掉内含子部分,然后再将外显子部分拼接起来。 - 5′末端加“帽”
-真核细胞成熟mRNA的5′末端均有一个特殊的结构,即m7Gpp-pmnNp,称为“帽”。 - 3′末端加“尾”
mRNA前体分子的3′末端有一段保守序列,由特异的核酸内切酶切去多余的核苷酸,然后在多聚A聚合酶的催化下,由ATP聚合生成多聚A尾。 - 碱基修饰
mRNA分子中有少量稀有碱基(如甲基化碱基)是在转录后经化学修饰(如甲基化)而形成的。 - 选择性加工
某些MRNA前体含有多个3‘剪切位点和多聚腺苷酸化位点,因此利用这些选择性位点可产生具有不同3’端非编码区或者具有不同编码能力的RNA产物。通过可变剪接途径可以挑先最保留在MRNA中的外显子,结果单个基因可以合成多种不同的蛋白质。 - RNA编辑
在合成并经RNA编辑加工之后,MRNA分子的序列可以发生改变。个别核苷酸可以被置换,添加或者删除。编辑过的MRNA翻译产生了较短脱脂基蛋白B48,由于基缺少一个结合受体的蛋白结构域,因此功能受限。还有好多其他编辑的例子,阵锥虫线粒体MRNA发生RNA编辑,使得最终MRNA中一半以上的尿嘧啶都获自编辑过程。
- 翻译
- 以mRNA 为模板指导蛋白质合成的过程
- 蛋白质合成在细胞中核糖体上进行
- mRNA tRNA和核糖体在翻译中起重要作用
- mRNA携带遗传信息
- tRNA转运活化的氨基酸和识别mRNA分子上的遗传密码
- 核糖体为蛋白质合成场所 把各种特定氨基酸分子连接成多肽链
- 翻译过程包括
- 起始
- 延伸
- 终止 - mRNA只决定氨基酸序列 蛋白质的空间结构是由翻译后修饰所决定的
RNA编辑
- 可改变初始转录物的编码特性
- 编辑的形式主要包括:
- 尿嘧啶核苷酸U的加入或删除
- C-U ,A-G或G-A的碱基转换
- C-G ,G-C或U-A的碱基颠换 - 编辑从mRNA的3-5 方向进行
- RNA编辑多种形式 极大增加mRNA的遗传信息量
