编译:微科盟小木,编辑:微科盟居居、江舜尧。
微科盟原创微文,欢迎转发转载,转载须注明来源《微生态》公众号。
导读
侵袭性真菌感染对公共健康构成了重大威胁,是抗菌素耐药性的一个未得到充分认识的组成部分,是全球新出现的危机。在全球环境发生深刻变化和高危人群不断扩大的时期,人类感染病原真菌正在对所有已获许可的系统性抗真菌药物产生耐药性。本综述重点介绍了抗真菌耐药性的主要机制,并探讨了细菌和真菌对抗菌素控制的耐药性之间的异同。讨论了旨在最大限度地减少病原真菌耐药性出现的风险降低策略所需的研究和创新主题。这些主题包括环境与全健康(One Health)之间的联系、监测、诊断、传播途径、新疗法和减轻真菌适应热点的方法。我们强调需要全球的努力来管理现有的抗真菌武器,并指导未来疗法和干预措施的研究和开发。
论文ID
原名:Tackling the emerging threat of antifungal resistance to human health
译名:应对新出现的抗真菌耐药性对人类健康的威胁
期刊:Nature Reviews Microbiology
IF:60.633
发表时间:2022.3
通讯作者:Matthew C. Fisher & Paul E. Verweij
通讯作者单位:英国伦敦帝国理工学院&荷兰国家公共卫生及环境研究院(RIVM)
DOI号:10.1038/s41579-022-00720-1
综述目录
1 前言
2 环境-全健康联系和新出现的抗真菌耐药性
2.1 唑类药物在环境和临床中的双重用途
2.2 不断变化的环境中耐药性对物种的影响
3 诊断和监测
3.1 鉴定抗真菌耐药性
3.2 耐药性检测、传播和监测
3.3 迈向抗真菌耐药性的全球监测
4 应对抗真菌耐药性的治疗方法
4.1 培育新的治疗方向
5 当前和未来的干预措施
6 结论
主要内容
1 前言
真菌在人类中引起多种疾病,从过敏综合征到浅表性、毁容和危及生命的侵袭性真菌疾病(IFDs),这些疾病共同影响着全球10亿多人。从历史来看,治疗主要依赖于四类具有全身作用的抗真菌药物:多烯类、唑类、棘白菌素和嘧啶类似物5-氟胞嘧啶,但真菌对化学攻击反应灵敏,治疗失败是常见的结果。这种失败归因于潜在的宿主免疫缺陷、抗真菌药物特性(药代动力学、药效学和药物-药物相互作用)和真菌特性(包括不同的细胞形态、抗真菌耐药性和抗真菌耐受性)之间的相互作用。对抗真菌药物的耐药性是世界范围内(在空间和时间上)新出现的问题,包括以前易感病原体的新耐药变种(如无处不在的霉菌烟曲霉Aspergillus fumigatus)以及对多种抗真菌药物具有耐药性的新物种(如耳念珠菌Candida auris)。这两种病原菌被列入美国疾病控制与预防中心(US CDC)于2019年发布的紧急抗菌素耐药性(AMR)威胁清单,现已正式认识到日益增加的公共卫生负担。
传统上,抗菌素耐药性计划排除抗真菌药物,因为作为公共卫生的威胁真菌被广泛忽视。真菌(真核)和细菌(原核)病原体之间的生物学差异也使真菌与现有AMR计划的整合变得复杂。然而,新出现的AMR问题在生命各个领域都存在,且耐药微生物之间存在许多相似之处(表1)。广谱抗菌抗生素(例如β-内酰胺类、头孢菌素类、碳青霉烯类、喹诺酮类和大环内酯类)的广泛使用通过清除易感基因型,使之有利于那些具有多态性和赋予耐药性的基因,从而深刻影响细菌群落,其中最适合的例子可以继续在全球范围内广泛传播。尽管研究还不够深入,但这一进化过程的各个方面都反映在整个真菌界,所有病原真菌都可以通过适应药物选择压力而获得耐药性。
从机制上讲,抗真菌耐药性通常是由于直接或间接影响药物靶点相互作用的变化而获得的。因此,耐药性可能通过靶结合位点的遗传变化(例如,编码唑类的羊毛甾醇脱甲基酶或编码棘白菌素的β-葡聚糖合酶的基因突变),通过过表达可用的靶标量和/或通过改变有效药物浓度(通过提高细胞内药物如唑类药物的药物外排活性,或抑制氟胞嘧啶的药物前体活化)而产生。概括的真菌耐药性机制见图1和BOX 1。与抗真菌耐药性相比,抗真菌耐受性是指药物敏感细胞在高于最低抑制浓度(MIC)的药物浓度下生长的能力,涉及广泛的一般应激反应和/或表观遗传途径。耐受性对于抗真菌药物最为明显,并且在用氟康唑治疗的白色念珠菌(Candida albicans)分离株中得到了最广泛的测定和表征。然而,其临床重要性仍然是一个悬而未决的问题。
抗真菌药物耐药性的获得和出现从根本上说是对药物施加的选择压力的进化反应。由于遗传变化而出现耐药性的可能性取决于暴露于选择压力的种群规模、细胞倍增率、赋予耐药性的不同途径(生理机制和因果遗传变化)的数量以及与每种途径相关的适应性成本。重要的是,抗真菌药物耐药性可能起源于宿主或环境。一方面,在抗真菌治疗过程中,个体体内耐药性从头开始进化,导致包括霉菌和酵母菌在内的一系列致病真菌的治疗失败。这与多种念珠菌(Candida)高度相关,它们是医院血流感染的主要原因,并显示出对抗真菌药物的广泛耐药性。例如,在HIV感染者长期使用氟康唑治疗口腔念珠菌病的过程中,白色念珠菌(C. albicans)对唑类药物产生耐药性已得到充分证明。这种现象并不局限于唑类抗真菌药物,因为在长期使用卡泊芬净治疗白色念珠菌食炎期间,也有报道称棘白菌素活性逐渐丧失。另一方面,由于人类病原真菌事先暴露于自然界中的杀菌剂,可能会产生环境抗性。杀菌剂的应用取决于长期需要保护集中养殖的动物和种植单一的、遗传同质的作物免受真菌感染,以及保护材料免受真菌的腐生腐烂。杀菌剂的环境压力推动了对所有主要杀菌剂类别的耐药性的演变,包括苯并咪唑、苯胺嘧啶、嗜球果伞素、琥珀酸脱氢酶抑制剂和包括唑类在内的甾醇脱甲基化抑制剂(DMIs)。环境抗性不仅需要制定抗性管理策略和培育更具抗病性的作物,它还与在环境和临床中使用甾醇14α-DMIs导致人类出现抗真菌IFDs有着密切的联系。耐药性真菌在自然界和临床中的出现以及风险患者群体的不断扩大,促使国际资助机构将抗真菌耐药性纳入其研究议程。值得注意的是,抗菌素耐药性联合规划倡议(JPIAMR)联盟于2021年4月首次将抗真菌药物耐药性纳入其抗菌素耐药性战略研究和创新议程。其综合的“One Health”框架整合了应对抗真菌耐药性的六个优先主题:环境、传播、监测、诊断、治疗和潜在干预措施。本篇综述中,我们将重点放在这些优先领域,旨在概述当前和未来的战略,以及应对抗真菌耐药性这一新兴公共卫生问题所需的关键研究。
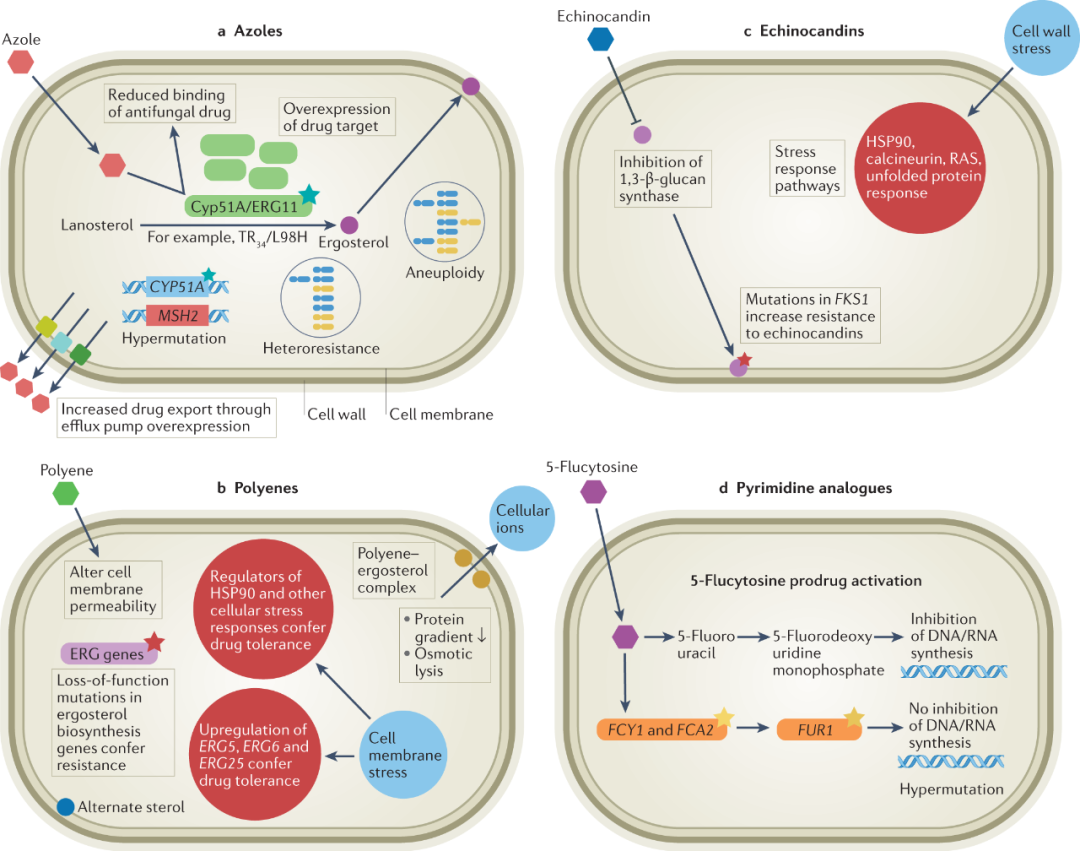
图1 主要侵袭性人类病原真菌获得抗真菌药物耐药性和/或耐受性的主要途径。获得抗真菌药物耐药性和/或耐受性的途径因作用方式(MOA)而异。a唑类药物耐药性主要是由于药物从真菌细胞外排的增加(特别是念珠菌)以及由CYP51A(烟曲霉)点突变和启动子插入引起的甾醇生物合成途径的修饰。在其他真菌物种中(如Cryptococcus neoformans),由染色体非整倍体和超突变引起的药物靶标和外排泵的过度表达是常见的。b多烯通过与麦角甾醇形成复合物来改变细胞膜通透性,而耐药性是由麦角甾醇生物合成基因的功能缺失突变引起的(特别是在曲霉属和念珠菌属)。特别是在白色念珠菌(Candida albicans)中,ERG3的双重缺失赋予了耐药性。然而,通过白色念珠菌中ERG5、ERG6和ERG25的上调,药物耐受性是常见的。c细胞膜应激也会影响HSP90的调节剂,从而赋予药物耐受性。棘白菌素抑制1,3-β-D-葡聚糖合酶(FKS1),该基因的突变导致念珠菌和镰刀菌的耐药性。棘白菌素暴露也可通过抑制β-葡聚糖合酶导致细胞壁应激,间接激活与药物耐受有关的Ca2+/钙调神经磷酸酶或HSP90/mTOR通路。d嘧啶类似物如5-氟胞嘧啶可抑制DNA和RNA合成。耐药性可通过靶基因FCY1的点突变而产生,并且在念珠菌属中很常见。已知隐球菌属物种中的超突变会引起对此类药物的耐药性。TR,串联重复。
BOX 1耐药性和耐受性的分子机制。
抗真菌耐药性被定义为在抑制生长和/或杀死该物种的大多数分离株的抗真菌药物浓度下生长的能力。一些物种对某些抗真菌药物具有内在抗性,这是由于与药物靶标的结合无效和/或在某一特定物种的所有成员中观察到的外排活性。例如,所有Aspergillus spp.、Candida krusei和大多数Candida auris分离株对氟康唑具有内在抗性,许多环境霉菌(如毛霉菌门Mucoromycota、Lomentospora spp.和镰刀菌Fusarium spp.)对唑类药物具有耐药性。获得性耐药是指获得耐药机制,使真菌细胞能够在比野生型种群成员更高的抗真菌药物浓度下生长。抗真菌耐受性,也称为拖尾生长或异源抗性,是指来自易感分离株的细胞亚群在药物浓度高于最低抑菌浓度(MICs)的情况下生长的能力,尽管生长缓慢。耐受性被认为是通过遗传、生理或表观遗传对药物的适应而产生的,遗传背景会影响表现出耐受性生长的潜力。抗真菌耐药性和抗真菌耐受性这两个术语在文献中经常被错误地交替出现。抗真菌耐受性的定义不同于抗菌素耐受性和持久性的定义,其中几乎所有细胞或非常罕见的细胞分别通过不同持续时间的短暂代谢静止而在杀菌药物治疗中存活。
可导致耐药性且真菌和细菌常见的突变类型(表1),包括点突变(每代每个细胞约10-6-10-8个)、基因复制和转座子插入(每代每个细胞约10-3-10-4个)。与细菌不同,真菌通常是多核和/或多细胞的,携带多条染色体。这种基因组组织为促进适应和耐药性出现的基因变化提供了更多机会(图1)。例如,临床相关的对唑类的耐药性和/或耐受性可以通过不同类型的突变演变,包括全染色体和节段性非整倍体。二倍体生物杂合性的丧失可以通过药物应激选择不同的杂合性丧失事件来增加耐药性或耐受性。抗真菌耐药性的发生也可能是由于光滑念珠菌和隐球菌属物种中的超突变真菌谱系所致,尽管这些特定突变在多大程度上导致突变率升高仍然难以捉摸。已知的真菌超突变状态的驱动机制集中在DNA错配修复机制上,主要通过MSH2突变产生,这些突变是通过对药物暴露的快速宿主适应或在致病真菌的自然谱系中发生。与经常遭受严重适应缺陷的细菌超突变谱系不同,真菌超突变谱系的适应成本并不高。
从不同个体分离的真菌基因型对唑类药物的耐受性水平差异很大,这可能是由于分离株之间的全基因组单核苷酸多态性(SNPs)具有相当大的多样性。在抗真菌药物暴露期间,药物耐受性的变化出现的频率高于耐药性水平的变化。据推测,突变后导致耐受性的途径数量大于直接影响耐药性的基因数量。在选择过程中,增加耐受性的突变很可能也会增加耐药率。与细菌一样,这可能是由具有获得和修复耐药性突变潜力的细胞群有效大小的增加所驱动的。
唑类药物通常对真菌有抑制作用,一般需要长期服用,而对多烯类药物(如两性霉素B)的耐药性相对较少出现,在临床上也很少见到。这可能是因为两性霉素B与麦角甾醇结合,与蛋白质靶标不同,麦角甾醇不是基因编码的。当出现多烯类耐药性时,似乎是由于通过消耗或替换麦角甾醇来调节细胞膜组成。
表型异质性可能会改变抗真菌药物的敏感性。例如,生物膜形成,一种多态细胞的固定生理状态,是耐药性和/或耐受性的非遗传途径。生物膜中的真菌细胞产生细胞外基质,作为药物汇,降低生物膜内细胞的有效药物浓度。此外,由瞬时遗传过程(如染色质修饰)维持的表观遗传状态可能会影响耐药性和/或耐受性,这可以通过组蛋白去乙酰酶抑制剂在体外突变时改变抗真菌药物反应得到证明。
2 环境-全健康联系和新出现的抗真菌耐药性
机会性致病真菌常见于我们周围的生活环境中,许多可以产生大量的空气传播孢子。因此,人类每天暴露于生物气溶胶等各种环境真菌病原体中。虽然大多数环境真菌在健康个体中不会引起明显的病理生理事件,但那些健康或免疫力受损的人容易感染一系列疾病,包括浅表性、过敏性、慢性和危及生命的IFDs。面临IFDs风险的患者群体目前正在扩大,并且(值得注意)包括老年人、免疫系统受到HIV、癌症化疗或移植所需的免疫抑制治疗影响的人,以及患有流感病毒和COVID-19等严重病毒感染的人。后一组患者的真菌感染激增,特别是曲霉属(Aspergillus spp.)、念珠菌属(Candida spp.,如耳念珠菌)和印度的毛霉菌门(Mucoromycota)物种对抗真菌治疗表现出强大的内在抗性和获得性耐药性。
分子流行病学研究一再表明,许多真菌疾病是通过我们周围的环境获得的;对于由球孢子菌属(Coccidioides spp.)、烟曲霉(A. fumigatus)和隐球菌属(Cryptococcus spp.)物种引起的IFDs尤其如此。真菌的环境种群与随后的抗真菌药物暴露之间的密切关系意味着新出现的环境抗性可能会影响真菌感染的临床管理。在农业环境中,植物病原真菌不断进化出对针对它们的一系列杀菌剂的耐药性。这种快速适应需要一个持续的发展周期,因为农业综合企业需要合成现有杀菌剂的变体或开发新的化学物质来阻止耐药性的积累。然而,与获得许可的医用抗真菌药物一样,农业中使用的农业杀菌剂在真菌界具有广谱活性。因此,耐药性不仅出现在作物病原体本身,也出现在包括潜在人类真菌病原体在内的其他环境真菌中。
广泛使用广谱农业杀菌剂对全健康的影响已针对DMI唑类药物进行了深入研究,其中这些化合物(如苯醚甲环唑、氧环唑、丙环唑和戊唑醇)不仅在结构上与一线医用三唑类化合物相似(艾沙康唑、伊曲康唑、泊沙康唑和伏立康唑),但在世界范围内的使用量越来越大。从2006年到2016年,美国唑类杀菌剂的使用量增加了400%以上,达到每年约3000公吨。中国的使用量是欧盟的十倍(每年约30000公吨),欧盟也有类似的情况。唑类杀菌剂的降解半衰期较长,从戊唑醇的47天到氧环唑的120天不等。鉴于其每年在全球的使用,预计环境中存在大量的唑类持久性,并有可能促进机会性真菌的耐药性或耐受性。自20世纪80年代唑类药物被广泛使用以来,环境和临床上对唑类药物产生耐药性的人类真菌病原体在世界范围内的增长情况已被记录下来,这是将农业杀菌剂的使用与临床中迅速增长的耐药性联系起来的确凿证据。
表1 比较细菌和真菌抗菌素耐药性的驱动因素。
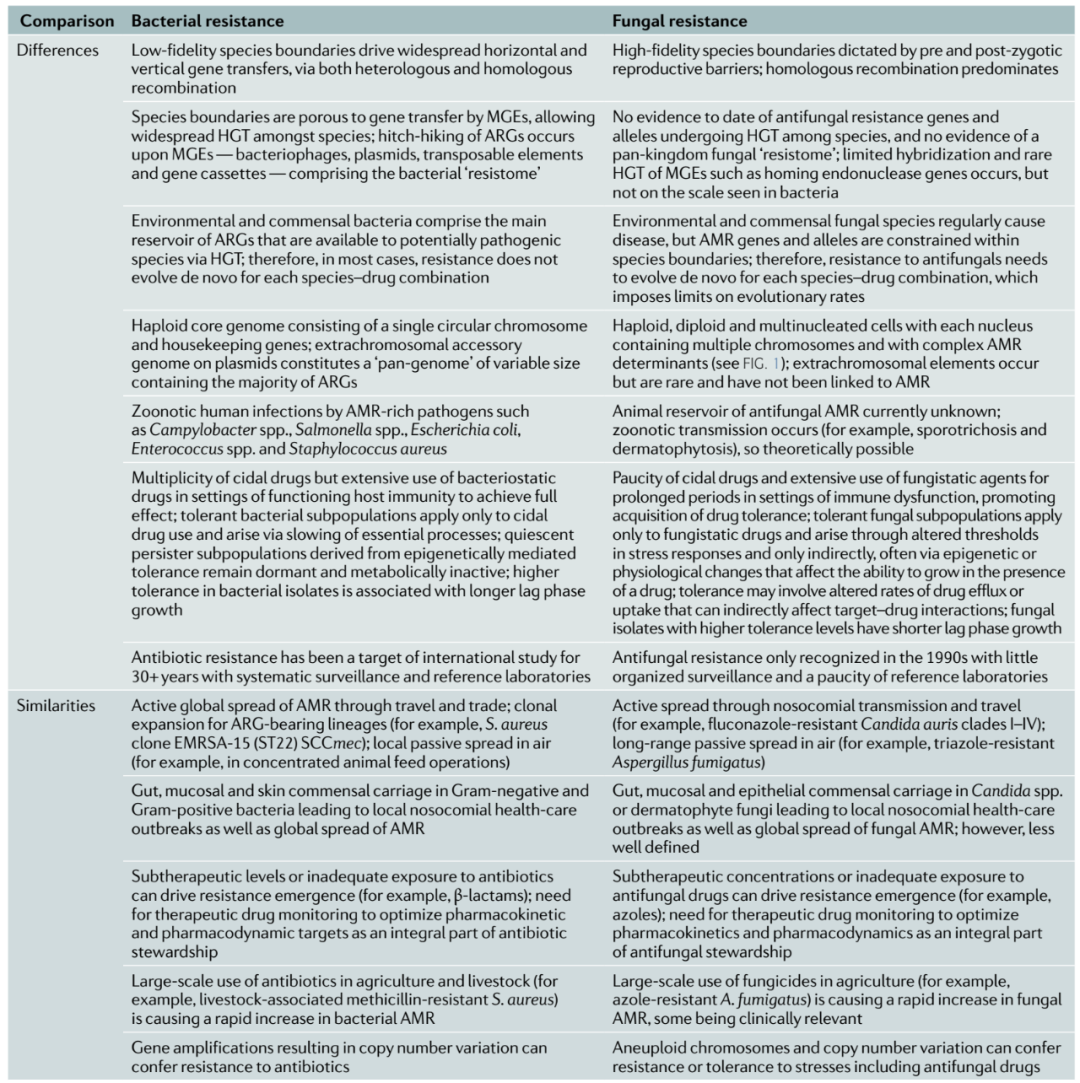
AMR,抗菌素耐药性;ARG,抗菌素耐药基因;HGT,水平基因转移;MGE,可移动遗传元件。
2.1 唑类药物在环境和临床中的双重用途
在环境中和以前没有抗真菌治疗史的患者中出现唑类耐药性烟曲霉的初步报告后,已经广泛探索了烟曲霉的环境耐药性和临床耐药性之间潜在的生态进化联系。假设了生态热点,即生物和非生物条件将趋同,允许真菌在接触亚MIC浓度唑类药物时生长,从而产生适合适应药物压力的条件(图2)。对这一假设的支持来自对在农业DMIs存在下支持烟曲霉高生长率的环境的研究;这些环境包括家庭和工业堆肥厂、城市环境和温室。
烟曲霉对三唑类药物的环境耐药性特征在于标志性遗传变化,包括CYP51A启动子区域中表达上调串联重复序列(TRs),其驱动基因表达增加,伴随着改变药物靶点的基因内点突变(图1)。临床内外最常见的耐药等位基因TR34/L98H和TR46/Y121F/T289A分别与伊曲康唑和伏立康唑耐药性相关,并且在世界范围内越来越多地被发现。分子流行病学方法发现了许多来自环境和受感染个体的成对耐药分离株的示例,具有统计学意义的遗传同一性,这意味着感染源是耐药环境分离株。因此,热点问题不再是环境中的耐药性会影响患者吗?相反,耐药性在哪里以及为什么会演变?它是如何扩散的以及可以采取哪些措施来减轻它?已成为关键问题。已证明耐三唑类烟曲霉有可能通过园艺产品(如交易植物鳞茎)在全球传播,并可加以管制。然而,分生孢子在气流中的传播是不可控的。此外,虽然人类并未被广泛视为耐唑类烟曲霉的生态相关来源,但某些患者群体在卫生保健机构中获得和传播耐唑类病原体的可能性意味着不能将他们排除在耐药接种源之外。
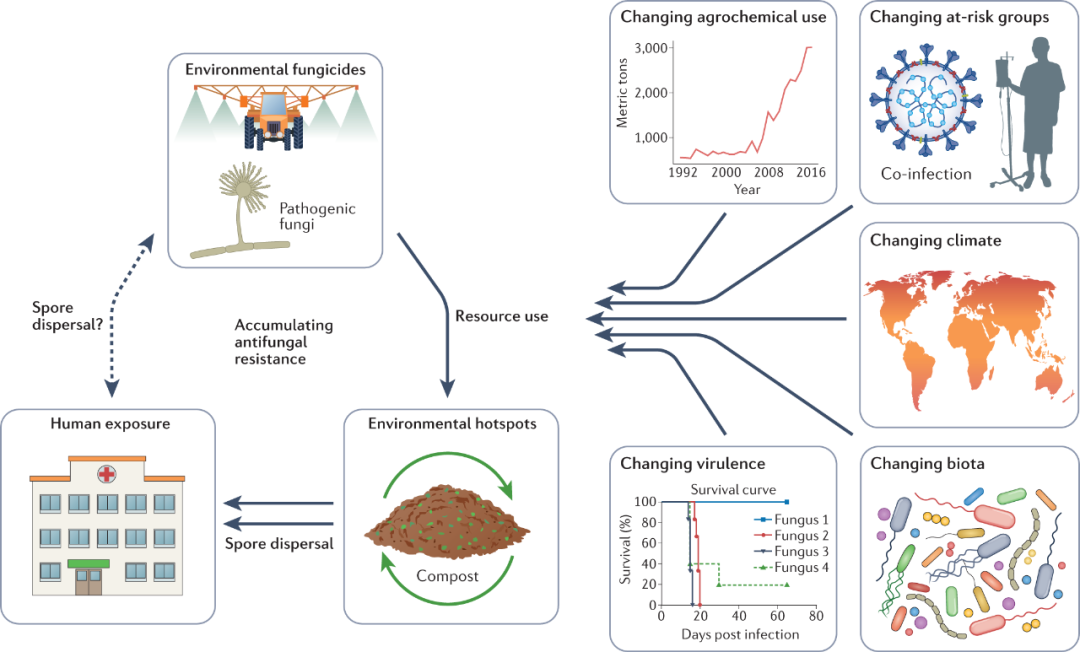
图2 新出现的抗真菌耐药性和环境-全健康驱动因素。环境中的真菌暴露于广谱抗真菌药物,这些药物也被用作临床抗真菌治疗的一线药物。生态热点的出现可以作为耐药基因型的扩增器。一个例子是绿色废物的储存和堆肥。患有侵袭性真菌疾病(IFDs)的人也可能传播耐药基因型(例如在医院内暴发);然而,人类和其他动物对环境中抗真菌耐药性的影响程度仍然未知。多种外部因素可能会影响抗真菌耐药性发生率,其中包括改变环境和废物管理中杀菌剂的使用模式;改变有风险的人类宿主群体,包括COVID-19等病毒感染;气候变化可能会改变真菌的地理范围和适应性景观,从而产生耐药性,并提供新的感染途径(例如自然灾害);改变可能包括类似于抗真菌药物的外源化学物质的生物相互作用;以及由于内在的遗传变化或与上述驱动因素组合的协同作用,真菌本身的毒力发生变化。
2.2 不断变化的环境中耐药性对物种的影响
环境杀菌剂的选择可能对人类真菌病原体的种群遗传结构及其遗传编码的表型性状产生广泛影响。烟曲霉中TR34/L98H抗性性状的出现与携带该等位基因的特定唑类耐药克隆的频率不断增加有关。然而,对烟曲霉基因组的扫描表明,唑类选择会导致在多个基因组区域和特定遗传背景下的选择性扫描。因此,对环境中杀菌剂的适应可能导致表型变化超过耐药性机制编码的变化。其中一个例子涉及到一个假说,即对唑类药物的耐药性也可以促使烟曲霉适应感染相关压力和毒力。甾醇生物合成(唑类的分子靶标),铁稳态和氧传感密不可分,因为麦角甾醇的生产需要许多铁依赖性酶,并且高度依赖氧。由于宿主环境是铁和氧的限制性环境,因此烟曲霉基因组中通过增强铁吸收和适应缺氧来增加唑类耐药性的任何变化都有可能同时促进毒力增强,这一假设应该得到验证。同样,格特隐球菌(Cryptococcus gattii)对广谱杀菌剂苯菌灵的适应与对氟康唑的交叉耐药性和小鼠毒力增加有关,这是一种归因于MDR1外排泵过表达的表型。在另一个例子中,在气候变化情景下预期的较高平均温度可能会影响抗真菌耐药性的出现。真菌通过调节细胞膜脂质组成来响应温度,例如,通过调节麦角甾醇生物合成途径,从而间接改变抗真菌耐药性。在堆肥、温室和热带国家等高温环境中,耐唑类烟曲霉的频率升高,这表明温度和抗真菌耐药性之间确实存在协同作用,但需要进一步的研究来确定这些相互作用的方向性和重要性。与此同时,温度(对气候变暖的热适应)和杀菌剂暴露之间的协同作用被用来解释在2009年发现的耐多药耳念珠菌在全球范围内迅速出现的原因。
关于真菌适应农用化学品后的遗传结构和适应性景观以及这如何影响它们与环境变化其他方面的相互作用,仍有很多需要研究的地方(图2)。因此,应对抗真菌耐药性的全健康解决方案必须跨越特定地点的地方(如含有农业化学残留物的绿色废物堆肥)和全球(如贸易和气候变化中的生物安全)规模。耐药性的演变可能导致更广泛的表型变化(如毒力增强),这可能是初始突变的直接结果,也可能是对患者或农业环境中富含唑类环境的二次适应。适应性的变化可能最终会影响停药后它们的持久性,未来的研究应该包括评估停止使用或从未使用过唑类药物的样本点的耐药基因型的“背景”频率。这些复杂的生态进化情景提高了了解抗真菌耐药性对真菌病原体、其生态学以及我们暴露于此类生物体的结果的全健康后果的必要性:要了解这一点,就需要加强监测。
3 诊断和监测
3.1 鉴定抗真菌耐药性
抗真菌耐药性(和耐受性)的鉴定依赖于培养微生物的药敏试验,确定特定抗菌素的MICs,与临床断点相比,这些抗菌素定义了易感性或耐药性。有几种方法可用于抗真菌药敏试验:肉汤微量稀释法、圆盘扩散法、唑琼脂筛选法、梯度扩散法和使用快速自动化仪器。临床和实验室标准协会(CLSI)和欧洲抗菌药物敏感性试验委员会(EUCAST)组织制定了进行敏感性检测的标准,并确定了有效治疗感染的“临床断点”。然而,标准化的CLSI和EUCAST肉汤微量稀释参考方法作为抗真菌药敏试验的金标准是劳动密集型、耗时的,并且在大多数临床实验室中很少进行。此外,它们需要从临床标本中进行真菌培养,这限制了灵敏度并且检测不到不可培养的Pneumocystis jirovecii。仅针对最常见物种(如白色念珠菌、光滑念珠菌Candida glabrata、热带念珠菌Candidatropicalis、Candida parapsilosis和烟曲霉)的主要抗真菌药物定义了临床断点,并且过度依赖这些作为研究较少物种的代理断点。EUCAST和CLSI断点之间的巨大差异使比较更加复杂。断点的应用依赖于准确的物种水平识别;随着越来越多地使用MALDI-TOF质谱系统,酵母菌的这种情况得到了改善,但霉菌仍然依赖于本地数据库内容。利用MALDI-TOF平台直接检测酵母和霉菌的抗真菌耐药性是一个新方向,但MALDI-TOF对许多中心而言成本太高(从而使国际耐药性监测计划复杂化),并且依赖培养增加了诊断时间。
3.2 耐药性检测、传播和监测
分子诊断方法具有识别可能与抗真菌耐药性相关的遗传标记以及识别具有内在抗性的真菌物种的能力已证实,但还未充分利用;其敏感性允许直接应用于临床标本,避免了培养的需要并缩短了周转时间。烟曲霉复合体的物种(如Aspergillus lentulus和Aspergillus felis)难以用常规方法进行鉴别,且对唑类抗真菌药物的MIC值可能较高,可以通过实时PCR进行鉴别。耐药念珠菌属物种,如耳念珠菌、光滑念珠菌(C. glabrata)和克鲁斯氏念珠菌(Candida krusei),可以通过PCR检测和鉴别,可能有助于感染控制和患者管理。全自动分子平台(T2 Biosystems或Becton Dickinson Max)的使用提供了快速检测系统,只需最少的专业培训,可与用于检测耐多药结核病的Cepheid GeneXpert平台相媲美。然而,必须扩大这种潜在的接近患者检测的范围,以包括检测与一般易感真菌物种的耐药性相关的突变。
通常通过直接测序编码药物靶蛋白的基因(如烟曲霉中的CYP51A或念珠菌中的ERG11)来识别潜在的耐药性相关突变。随后,基于常见突变的高流行率(如烟曲霉中的TR34/L98H和TR46/Y121F/T289A以及P. jirovecii中的二氢蝶酸合酶突变),推出了商业实时PCR检测,并且由于基于PCR的方法的高灵敏度和特异性,它们的诊断用途正在增加。由于念珠菌(Candida spp.)对唑类药物的耐药性与多种机制和随后的突变有关,因此实时PCR方法的发展受到限制。DNA测序仍然是识别与唑类耐药相关突变的最佳选择,限制了临床应用,特别是直接样本检测。对唑类和棘白菌素耐药的耳念珠菌菌株ERG11和FKS1基因进行测序,发现了相关的热点和特定突变,为开发快速分子检测提供了条件。少数FKS1基因突变与念珠菌属(Candida spp.)物种中大多数对棘白菌素的耐药性有关,并且已经开发了PCR检测。目前,还没有商业化的PCR检测来检测与酵母抗真菌耐药性相关的突变,并且为了改善诊断,必须通过加强商业开发和监管机构的支持来认识到这一点。
技术和计算方法的进步促进了耐药性检测。此处的示例包括通过环介导等温扩增将无热循环仪的DNA扩增整合到带有硅芯片检测器和云连接的芯片实验室平台上,以实现未来的即时抗性检测或新开发的焦磷酸测序技术。全基因组测序(WGS)的实施为更全面地探索基因突变的生物学基础提供了广阔的前景。常规应用WGS进行细菌病原体鉴定、耐药等位基因检测和传播途径鉴定已成为普遍现象。除了检测耐药等位基因外,WGS的一个主要优势是能够重建AMR变异的时空进化轨迹。然而,与抗菌素耐药性相比,标准化的WGS分型方法并没有广泛应用于真菌,因为它们的基因组更大,有性重组频繁,缺乏标准化的生物信息学管道。对抗真菌耐药决定因素和物种基因组的进一步了解将支持向基于WGS的对几种人类真菌病原体的AMR的理解转变。为了实现这一目标,快速基因组分析的发展已经成为了解耳念珠菌(C. auris)在国际和地方范围传播的关键,包括多重耐药变体的出现。不可培养的真菌是一个挑战,需要更有针对性的方法。例如,针对P. jirovecii的多位点序列分型方案可实现抗真菌耐药性标志物分析。对于曲霉属物种(Aspergillus spp.),需要更多关于耐药性机制的知识,因为许多耐药菌株不携带少数已知的耐药相关等位基因。尽管如此,WGS越来越多地用于追踪烟曲霉中已知多态性的AMR传播。即时WGS设备(如纳米孔测序仪)能力的提高正在加快我们检测抗真菌耐药性突变的能力,并可能改变我们了解疫情暴发环境中医院传播途径的能力(图3)。
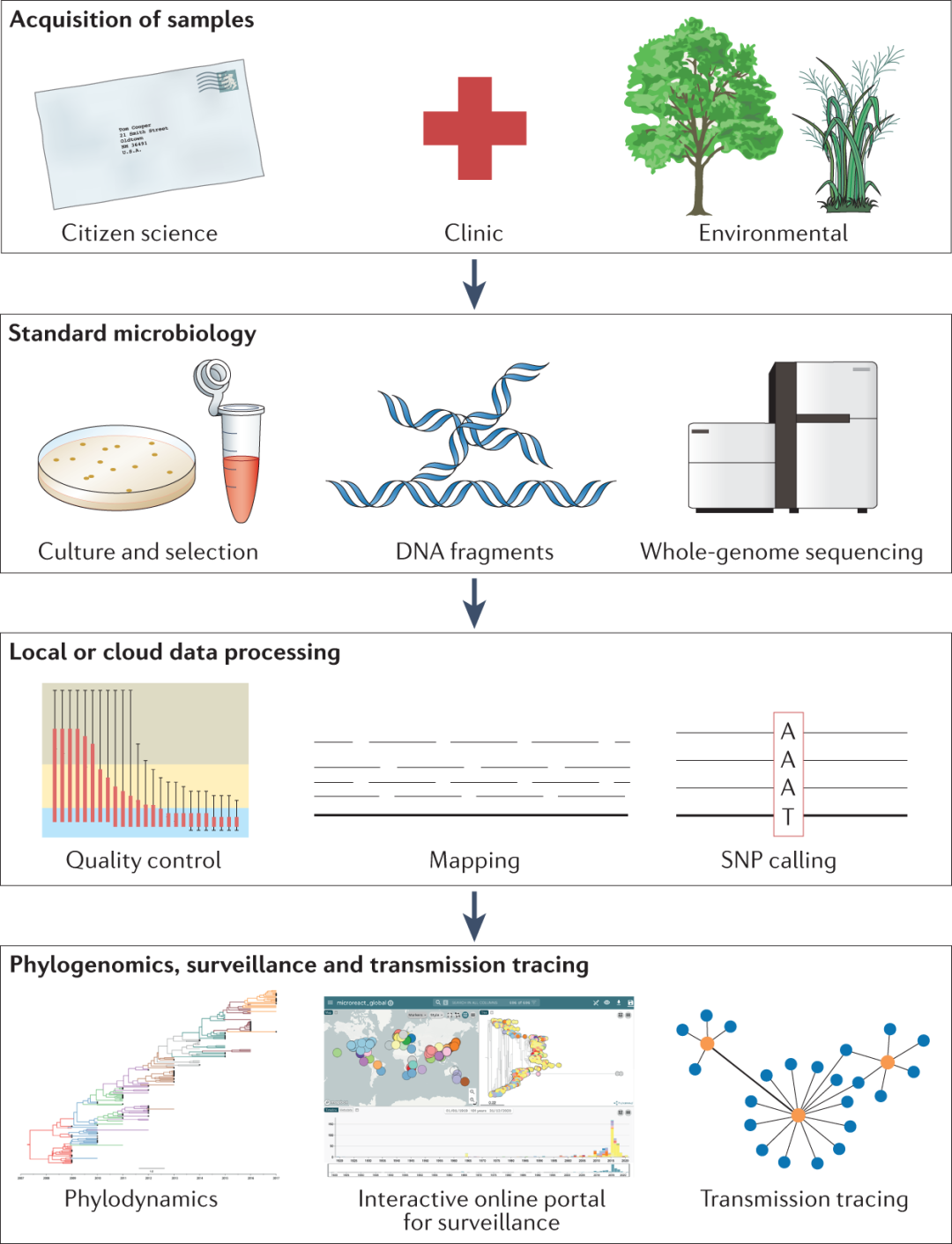
图3 耐药性检测、追踪和监测。真菌样本可以从诊所或环境中获取,包括以公民科学家的身份与公众接触。传统的、成熟的微生物学方法可以从这些样本中培养和选择分离株,为提取基因组DNA做好准备。这些DNA片段用于生成全基因组测序(WGS)的测序文库。目前有许多可用的测序平台,可生成长读长和短读长序列数据。在对参考基因组进行映射之前,需要对原始序列数据进行质量控制。调用高置信度单核苷酸多态性(SNPs)可以帮助推断与耐药性相关的等位基因及其进化历史。研究人员和临床医生可以通过系统动力学推断和建立交互式在线门户(如Nextstrain或Microreact)来追踪传播事件。
3.3 迈向抗真菌耐药性的全球监测
公共卫生机构已在许多国家发起了对细菌AMR的系统监测,并指定了参考实验室与常规医学微生物实验室联系。由美国疾病控制与预防中心(CDC)和欧洲疾病预防与控制中心牵头的大型国际监测研究对耐抗生素细菌的传播进行监测,并发布早期预警信号。然而,迄今为止,大多数AMR监测计划都将真菌排除在外。2018年,WHO(世界卫生组织)启动了一项念珠菌监测试点计划,以收集关于侵袭性念珠菌分离株抗真菌耐药性的回顾性数据;最近正式将其纳入全球抗菌素耐药性监测系统(GLASS)计划(方BOX2)。CDC的新发感染项目目前在美国10个州的卫生部门开展基于人群的积极监测,监测念珠菌血症的流行病学趋势。在全球范围内,SENTRY抗菌素监测计划至少有427个参与中心,并通过间接(通过血液培养监测)和直接收集抗真菌耐药性数据。不幸的是,提供真菌病原体数据的中心相对较少。除了这些更广泛和更系统的监测计划外,还可从澳大利亚、苏格兰、芬兰、冰岛、挪威、瑞典、英国和丹麦等几个国家获得对念珠菌的全国监测数据。然而,对其他真菌物种的监测很少见,大多数已发表的数据仅限于耐唑类烟曲霉。
抗真菌耐药率的上升和新的多重耐药物种(如耳念珠菌)在全球范围内的迅速出现使得将真菌感染纳入现有的国家和国际监测计划势在必行。尽管在世界范围内检测到烟曲霉的唑类耐药基因型,但在大多数临床环境中,没有对其进行检测,也很少有研究探讨其与临床失败的关系。值得注意的是,作为号召性用语,WHO目前正在定义一个与细菌病原体相一致的真菌病原体优先清单,这是可能引发该领域研究和创新的重要一步。当前的一个高度优先事项是需要通过收集基本临床和流行病学数据来实施标准化监测。这是因为改善监测将进一步增进对真菌AMR进化和传播的了解,同时有助于实施现代基因组监测方法。同时,迫切需要建立包括研究、临床和行业合作伙伴在内的协作网络,以开展多中心研究;这些网络还将需要访问共享的生物存储库,这些生物存储库将经过验证的样本与元数据进行整理,并在需要时快速、公平地分发这些数据。在需要对临床真菌学专业知识进行能力建设的高资源和资源有限国家,应在当地的临床实验室加强准确的真菌物种鉴定、简单的耐药性筛查方法和MIC检测。当在当地发现耐药菌株时,参考实验室的确认与基本临床和流行病学数据的收集相结合,将有助于下游制定政策建议和控制战略。
BOX 2 优化抗真菌耐药性监测的优先事项
现有的抗真菌监测
•SENTRY抗菌药物监测计划
•WHO(世界卫生组织)全球抗菌素耐药性监测系统(GLASS):Candida spp.。
•特设国家和地方规模的抗真菌监测
未来抗真菌监测的重点
•开发和调整适用于低收入和中等收入国家的工具,并建设使用这些工具的能力。
•增加适用于当地临床实验室的快速简单的抗真菌耐药性筛查技术的可用性。
•指定国家参考实验室来监测抗真菌耐药性。
•加强基础研究,以确定抗真菌耐药性的分子机制和相关诊断标志物。
•开发标准化临床研究数据库,将体外和体内耐药性与临床结果联系起来。
•创建全球可访问的重点人类真菌病原体基因组抗真菌耐药性数据库。
•在国家和国际层面实施标准化和关联的抗真菌耐药性监测网络,并采用国际统一的定义和数据类型。
•建立可访问的样本生物存储库和元数据,以加速学术和行业合作,开发耐药性诊断方法。
•在OneHealth范式下将人类抗真菌耐药性监测扩展到兽医、野生动物和环境样本。
4 应对抗真菌耐药性的治疗方法
对于共生微生物,接受治疗的个体可通过药物暴露获得抗真菌耐药性。例如,棘白菌素耐药性在之前接受过棘白菌素治疗的个体中更为常见,而Cryptococcus neoformans和A.fumigatus的唑类耐药基因型在长期治疗过程中出现。为了使抗真菌药物有效,它们必须到达感染部位。每一种抗真菌药物的吸收、分布、代谢和排泄(药代动力学)特性都有很大的差异,更明显的是药物在组织特异性渗透方面的差异。持续低或短暂高的药物浓度可能会加速耐药性的演变。然而,使用过高剂量的药物会带来伴随的毒性风险。由于这些原因,需要定期监测治疗药物以优化剂量,最大限度地发挥治疗潜力,并尽可能减少耐药性的演变,同时最大限度地减少不良反应。组织特异性药代动力学在很大程度上是未知的,尽管基于生理学的建模方法已经开始揭示这一问题。现实世界的研究越来越多地使用治疗药物监测来探索临床队列中的药代动力学,例如,对囊性纤维化患者的监测表明,亚治疗水平的唑类药物的流行率很高,并且在2年后发生耐药感染的可能性很高(>20%)。出于这些原因,需要在易感人群中通过抗真菌管理计划更好地实施治疗药物监测。同时,知情的药物组合应用可能会规避耐药性。例如,米卡芬净可抑制多种人类和真菌外排泵,因此当与唑类药物联合使用时可增强其细胞内滞留和疗效。
未来的研究将需要确定耐药性和耐受性机制出现的可能性。药理学方法允许模拟模型预测,例如,中空纤维模型使用可用的药代动力学数据来模拟抗微生物药物的人体药代动力学。此外,由于广泛坏死导致不良结局,在感染部位的药物输送仍然是一个挑战。对于感染部位药物渗透性较差的疾病,需要改进药效学模型来优化给药方案,防止治疗失败。总之,药代动力学/药效学结合方法可以促进对(非结合)药物浓度、病原体生长和杀伤动力学之间相互作用的综合动态研究,以确定最大限度地减少原位抗真菌耐药性演变的条件。
4.1 培育新的治疗方向
对于可能因双重用途而受到影响的有限类别药物的相关问题,一个明显的解决办法是加速药物开发。但这不是一个可以迅速实现的解决方案,因为从首次人体试验到批准一种新型抗感染药物大约需要5-7年的时间,而且可能需要花费数亿美元。如果考虑到早期开发成本,时间和成本就会高得多。例如,由巴西利制药公司和随后的阿斯特拉制药公司和辉瑞公司开发的Cresemba(艾沙康唑)开发项目历时13年,需要大约1亿美元的资金,后续审批费用约为3000万美元。虽然艾沙康唑比伏立康唑更广谱,如对毛霉菌目(Mucorales)物种的疗效,并且对侵袭性曲霉病患者同样有效,且药物相关不良事件也比伏立康唑少,但该药物在曲霉菌和念珠菌中仍显示出对其他唑类药物的交叉耐药性。药物研发公司F2GLtd正在开发olorofim,这是一种靶向二氢乳清酸脱氢酶的新作用模式(MOA)抗真菌药物,自1998年成立以来,该药物已进行了数轮投资,总投资超过2.13亿美元。F2GLtd确定初始化合物并开发其进入II期试验的总时间约为23年。虽然该药物对念珠菌无活性,但对曲霉属物种具有良好的活性,包括具有获得性唑类耐药性的分离株和其他难以治疗的霉菌,如Lomentospora prolificans。这些例子强调了与识别和开发一类新型抗真菌药物相关的投资和风险。
这些高成本和冗长的时间尺度对于开发治疗由抗真菌物种引起的IFDs的疗法有明显的意义,其中大多数是相对罕见的,不太可能提供显著的投资回报。治疗这类疾病的新疗法可能只会作为广谱抗真菌药物的辅助药物出现,而广谱抗真菌药物主要用于治疗更常见的真菌疾病。然后出现了一个关键问题,即什么市场规模足以使抗真菌开发项目可行。一个答案可能在于有前景的真菌细胞壁几丁质合酶抑制剂Nikkomycin Z的开发,该抑制剂在I期试验成功后停滞不前。开发商Valley Fever Solutions Inc.迄今未能获得进一步开发该化合物的投资。这很可能与尼可霉素Z活性范围有限有关,它对相对罕见的地方性真菌(如球孢子菌属Coccidioides spp.)最有效,而球孢子菌的感染患者人数仅为2.5万人左右,预计每年的销售峰值为1.3亿美元。尽管这些感染大部分发生在美国,但投资者认为这个市场规模太小,尽管尼可霉素Z得到了政府倡议的支持(如孤儿药指定和快速通道指定)以及与其他抗真菌药物联合使用的良好效果。
抗真菌药物管道正在经历大幅增长,这表明价值130亿美元的全球抗真菌药物市场正在鼓励开发精制的既存化合物以及具有广泛活性的新型MOA抗真菌药物。值得注意的是,Gwt1抑制剂fosmanogepix(辉瑞新收购)、(1→3)-β-D-葡聚糖合酶抑制剂ibrexafungerp (Scynexis)和olorofim(F2G Ltd)都是新的MOA抗真菌药物,它们将为治疗唑类耐药或棘白菌素耐药病原体提供机会(补充图1)。正在开发的其他新型MOA抗真菌药物具有细胞内靶点,因此可能对对现有药物类别具有耐药性的分离株有效。
除了全身给药外,目前正在探索将抗真菌药物运送到作用部位的新策略。Opelconazole(Pulmocide)是一种通过雾化给药的新剂型唑类药物,已在I期试验中评估可用于治疗侵袭性曲霉病。由于可以在肺中达到更高的药物浓度,局部应用可以克服烟曲霉的唑类耐药性。下一代抗真菌药物需要考虑抗感染药物的使用寿命与潜在的耐药性出现率之间的关系。因此,因此,应在尽可能早的开发阶段确定新抗真菌药物耐药性的进化风险,正如抗菌药物管道所倡导的那样。建议采用能更好地复制体内环境的慢性曲霉病和急性念珠菌病模型或体外系统来监测体内耐药性发展的潜力,包括目标生物体和感染部位及远端身体部位的共生真菌。将这些体内模型与药代动力学/药效学模型相结合,可以促进剂量研究,估计耐药性出现的可能性,并最大限度地减少耐药性、真菌持久性和耐受性的出现。
在农业和医学中使用同一类药物是曲霉属物种(Aspergillus spp.)环境耐药性的关键驱动因素。从农业中去除唑类物质并不实际可行,因为它将对全球粮食生产产生重大影响。然而,植物病原体对唑类药物的抗性正在农业环境中迅速出现。那么,考虑到全健康,抗真菌药物的发展前景如何?显然,农用杀菌剂和药用抗真菌剂的发展需要分开。在农业领域,这可以通过开发作物的综合疾病管理来实现,包括具有病原体抗性基因镶嵌的进化智能抗病作物以及例如基于RNA干扰的物种特异性新型抗真菌治疗方法的开发。那些侧重于对植物致病性至关重要但与人类不同的靶标的方法也可能导致控制真菌病原体的不同方法。为此,已经取得了重大的技术进步,通过结合功能基因组学和下一代测序来实现毒力决定因素的高通量鉴定。毫无疑问,为农业综合企业和制药企业加快开发多样化、差异化和封闭式抗真菌管道,不仅是开发新的杀真菌化合物的关键,也是解决未来几年不断发展的抗真菌耐药性的关键。
5 当前和未来的干预措施
我们如何才能遏制新出现的抗真菌耐药性浪潮?整合JPIAMR和WHO倡议的“支柱”将保护和增强我们治疗IFDs的能力(图4)。目前限制人类真菌病原体进化为化学控制的可用策略包括加强监测和抗真菌管理计划,这两者都需要改进对IFDs和抗真菌耐药性的诊断;最大限度地减少环境-临床双重使用抗真菌药物;并使用现有的许可药物优化弹性联合疗法。未来减轻抗真菌耐药性影响的策略主要需要用仅用于临床的新型抗真菌化合物治疗高危人群。这种个性化医疗方法应包括通过使用免疫疗法和/或针对IFDs的疫苗来解决使个体易患这些疾病的免疫力减弱的问题,从而降低获得性IFDs的风险。
广泛的预防性和经验性抗真菌药物处方治疗长期处于风险中的人群(例如囊性纤维化患者)、重症患者和血液肿瘤患者的疑似IFDs仍然是一个令人担忧的问题。需要有效的抗真菌管理以优化抗真菌药物的使用并保护有限的抗真菌药物库。这与高度传播的真菌感染尤其相关,如念珠菌属(Candida spp.)和皮肤感染毛癣菌属(Trichophyton spp.)。
在主要为单中心、历史性队列观察(非随机)研究中,抗真菌药物管理计划一直证明在措施方面有所改进,如及时和适当的抗真菌处方(指南驱动)、诊断和药物监测的使用以及减少抗真菌药物消费,降低抗真菌药物的选择压力和耐药性的发展。虽然这些研究并非旨在证明临床结局的改善,但抗真菌药物管理的实施对IFDs的发病率、住院时间和住院死亡率没有不利影响是重要的发现。抗真菌药物管理以及时和敏感的诊断为基础,尽管对各种先发制人的诊断与经验性抗真菌策略的回顾证实了先发制人策略的适用性,但最佳策略和限制尚未确定。未来工作的目标包括优化快速诊断策略,以实现“早开始-降级-早停止”抗真菌策略和更好的医院感染控制,以及证明抗真菌药物管理对住院患者队列或医院环境的抗真菌耐药率的影响。
联合抗菌治疗是预防各种细菌和病毒感染继发性AMR的既定有效策略。该原则于20世纪50年代在结核病治疗中确立,并在20世纪90年代在HIV治疗和最近在丙型肝炎病毒治疗中重复使用。两性霉素B和氟胞嘧啶(在两性霉素B不可用的情况下氟康唑联合氟胞嘧啶)的联合治疗是隐球菌病的既定护理标准。氟胞嘧啶和氟康唑联合使用可防止隐球菌脑膜炎患者在氟康唑单药治疗后产生氟康唑耐药真菌种群的选择。就原发性、环境衍生的抗真菌耐药性而言,联合治疗患者的效果可能有限,但联合治疗可以减少因原发性耐药而导致的治疗失败,并限制继发性临床抗真菌耐药性的发展。就抗菌效果而言,联合治疗可能是附加的或协同的,需要进一步的工作来进一步发挥其在各种危及生命的真菌感染中的潜力。
对于侵袭性曲霉病,体外和动物模型数据一致表明,唑类和棘白菌素类药物联合使用可增加对真菌的杀灭作用并提高存活率。在一项随机临床试验中,使用这种组合的患者死亡率为19%,而使用唑类单药治疗的患者死亡率为28%;虽然这项研究的规模有限,意味着生存获益没有达到传统的统计学意义,但所描述的方法是令人鼓舞的。动物模型表明联合治疗在耐唑类侵袭性曲霉病中有一定作用,但在进行充分有力的III期试验之前,需要在实验模型和II期临床研究中系统地探索已建立的和新的抗真菌药物的联合治疗。与机会性病原真菌相比,耳念珠菌可在重症监护室和其他医疗机构设施中持续存在和传播,导致严重和棘手的医院暴发。棘白菌素单药治疗常用于耳念珠菌的治疗,而耳念珠菌通常对氟康唑耐药。由于这种方法可能会促进耐多药分离株的进化和传播,因此必须系统地评估联合治疗策略以减轻这种现已全球化的真菌的风险。
其他保护现有抗真菌药物的方法包括利用宿主导向方法管理抗真菌药物耐药性,其中包括免疫疗法、真菌疫苗和真菌靶标抗体。由于IFDs在免疫缺陷的宿主中最为常见,因此针对宿主的免疫疗法已经在开发中,如重组细胞因子、单克隆抗体和真菌特异性工程T细胞。使用干扰素-γ预防和治疗慢性肉芽肿病患者的侵袭性曲霉病是第一个成功的宿主定向抗真菌免疫治疗。自那时起,已有一系列病例报告描述了TLR7激动剂咪喹莫特成功用于染色芽生菌病和粒细胞-巨噬细胞集落刺激因子(GM-CSF)治疗与CARD9缺陷相关的中枢神经系统念珠菌病。这些进展凸显了以宿主为导向的方法减轻抗真菌药物压力的潜力。此外,基于细胞的疗法(如树突状细胞转移和嵌合抗原受体(CAR)T细胞疗法)已在体外显示出可喜的结果,但需要在临床试验中进行评估。
免疫疗法与常规抗真菌疗法的结合也有前景。许多候选真菌疫苗已经在临床前环境中进行了研究,但只有白色念珠菌重组Als3蛋白疫苗在II期临床试验中显示出了良好的结果。推进抗真菌疫苗需要克服几个障碍,特别是真菌在人类共生功能体中普遍存在的特性,以及易患IFDs人群中预期的次优免疫反应。同样显示出希望的是抗体和真菌模式识别受体,它们可能靶向抗真菌药物进行病原体递送。Dectin-2与两性霉素B脂质体偶联的临床前研究在实验性肺曲霉病中显示出令人鼓舞的结果,并可能有助于降低宿主的抗真菌毒性。然而,尽管以宿主为导向的抗真菌策略,单独或与传统抗真菌药物联合使用前景广阔,但从实验室到临床试验的这些新策略的推进和融资将是未来十年的重大挑战。
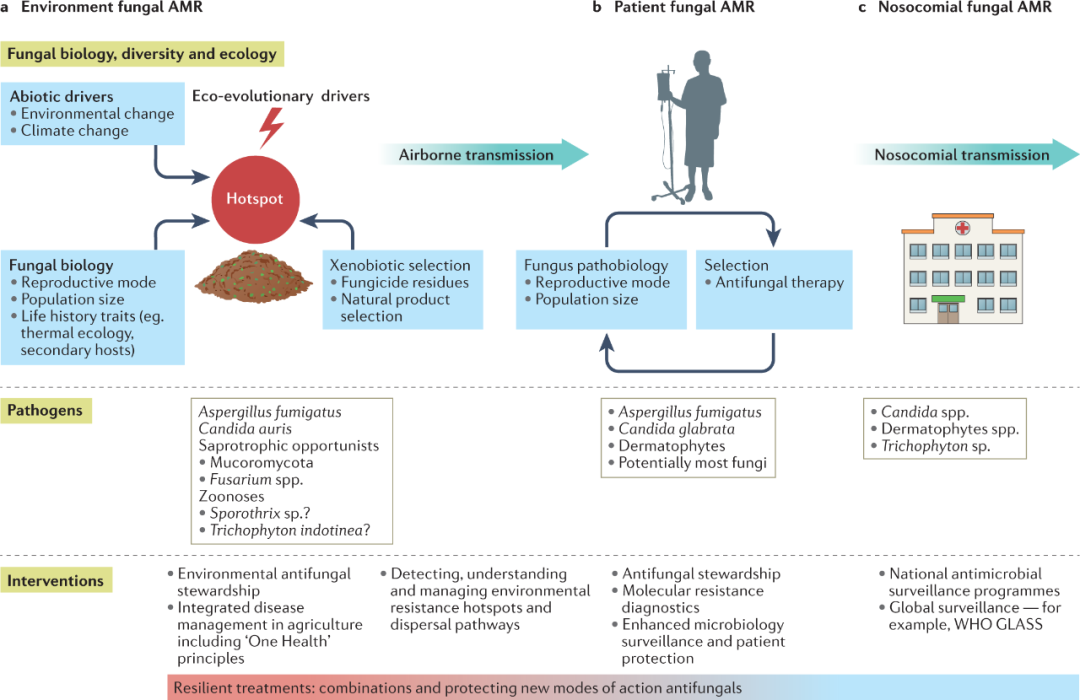 图4 在抗真菌耐药性的情况下对侵袭性真菌感染进行干预。对Synopticintegrated One Health的理解对于理解导致整个真菌界出现耐药性的复杂多因素途径以及降低出现率的潜在干预措施都是必要的。a复杂的生物和非生物相互作用导致环境机会性真菌出现AMR发展的进化热点,需要对环境进行有针对性的干预。b,c患者暴露于环境AMR需要加强检测方法,更多地关注关键的真菌生活史因素(b部分),而新的和正在出现的耐药病原真菌有可能在全球医院传播并在医疗保健环境中暴发,需要进行跨国监测(c部分)。一个跨领域的主题是,工业需要将农业杀菌剂的开发和使用与临床使用的抗真菌药物分开,以开发对a-c部分发挥作用的进化力量有弹性的治疗方法。GLASS,全球抗菌素耐药性监测系统;WHO,世界卫生组织。
图4 在抗真菌耐药性的情况下对侵袭性真菌感染进行干预。对Synopticintegrated One Health的理解对于理解导致整个真菌界出现耐药性的复杂多因素途径以及降低出现率的潜在干预措施都是必要的。a复杂的生物和非生物相互作用导致环境机会性真菌出现AMR发展的进化热点,需要对环境进行有针对性的干预。b,c患者暴露于环境AMR需要加强检测方法,更多地关注关键的真菌生活史因素(b部分),而新的和正在出现的耐药病原真菌有可能在全球医院传播并在医疗保健环境中暴发,需要进行跨国监测(c部分)。一个跨领域的主题是,工业需要将农业杀菌剂的开发和使用与临床使用的抗真菌药物分开,以开发对a-c部分发挥作用的进化力量有弹性的治疗方法。GLASS,全球抗菌素耐药性监测系统;WHO,世界卫生组织。
结论
当前临床医生管理耐药性IFDs的能力面临的挑战包括缺乏敏感性和特异性的诊断测试、缺乏经过临床校准的抗真菌药敏试验以及抗真菌药物种类有限。此外,真菌界的丰富度和多样性确保了新病原体的无底洞,以及源源不断的变种,当暴露于抗真菌化学物质时,这些变种很容易适应并进化。在快速变化的环境中,真菌物种的绝对生态广度及其独特而多样的生态营养意味着人类健康将始终与真菌群落的复杂生态学息息相关,无论是共生的还是环境的。同样,我们同时需要控制农业环境和临床中的真菌疾病,这意味着综合应对措施需要将这些需求考虑在内。病原真菌被广泛传播(包括主动和被动),因此在临床和实地处理抗真菌耐药性需要全球协调应对。目前缺乏对网络、基础设施、研究经费和职业发展的跨国支持,必须通过加强决策者、资助机构和研究人员之间的协调来解决,包括抗真菌药物的生产者和使用者。
原文链接:https://www.nature.com/articles/s41579-022-00720-1
猜你喜欢
10000+:菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature Cell专刊 肠道指挥大脑
文献阅读 热心肠 SemanticScholar Geenmedical
16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流、快速解决科研困难,我们建立了“宏基因组”专业讨论群,目前己有国内外5000+ 一线科研人员加入。参与讨论,获得专业解答,欢迎分享此文至朋友圈,并扫码加主编好友带你入群,务必备注“姓名-单位-研究方向-职称/年级”。PI请明示身份,另有海内外微生物相关PI群供大佬合作交流。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。

学习16S扩增子、宏基因组科研思路和分析实战,关注“宏基因组”
点击阅读原文,跳转最新文章目录阅读