点击蓝字 关注我们
玛咖来源细胞外囊泡通过脑-肠轴促进五羟色胺释放改善抑郁症

iMeta主页:http://www.imeta.science
研究论文
● 原文链接DOI: https://doi.org/10.1002/imt2.116
● 2023年6月4日,徐州医科大学齐素华团队和广东省人民医院顾兵团队等在 iMeta 在线发表了题为 “Lepidium meyenii Walp (Maca)-derived extracellular vesicles ameliorate depression by promoting 5-HT synthesis via the modulation of gut–brain axis” 的文章。
● 本研究结果表明玛咖来源的细胞外囊泡(Maca-EVs)可能通过调节肠-脑轴来促进五羟色胺(5-HT)的释放,从而改善抑郁行为。研究揭示了一种利用植物源细胞外囊泡治疗抑郁症的新方法。
● 第一作者:洪蕊、罗兰、王亮、胡兆丽
● 通讯作者:齐素华 ([email protected])、顾兵([email protected])
● 合作作者:尹麒龙、李明、顾斌、王斌、庄涛、张馨月、周愿、王婉、黄琳燕
● 主要单位:徐州医科大学医学技术学院、药学院、医学信息与工程学院、生物化学与分子生物学研究中心、江苏脑疾病生物信息重点实验室、附属医院检验医学科;广东省人民医院检验医学科;澳大利亚伊迪斯科文大学
亮 点
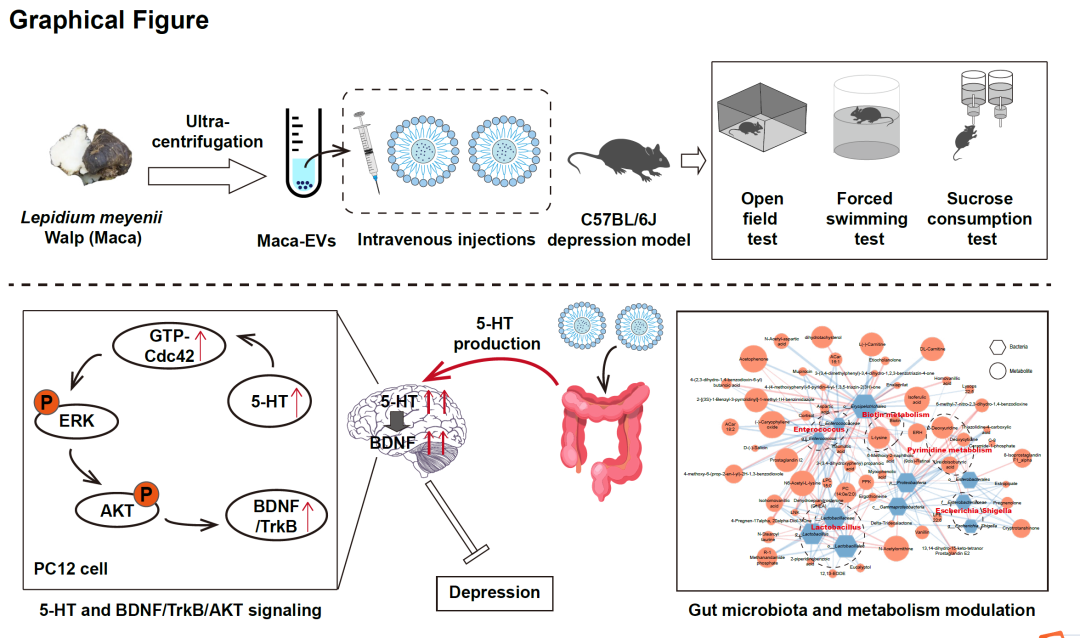
● 我们成功地使用超高速离心法分离了玛咖来源的细胞外囊泡(Maca-EVs),并在不可预测的慢性温和应激小鼠抑郁模型中确定其抗抑郁作用。
● Maca-EVs可能通过调节肠道微生物群和代谢增加五羟色胺(5-HT)水平。
● Maca-EVs的抗抑郁活性可能与血清5-HT水平升高以及GTP-Cdc42/ERK和BDNF/TrkB/AKT信号通路激活相关。
摘 要
抑郁症是一种常见的使人精神衰弱的疾病,需要有效的治疗。玛咖是一种具有治疗抑郁症潜在药理作用的植物。最近,由于植物来源的细胞外囊泡(PDEVs)具有低毒性和运输到人体细胞的能力,人们对其越来越感兴趣。靶向肠-脑轴是一种新的抑郁症治疗策略,可以通过使用玛咖来源的细胞外囊泡(Maca-EVs)来实现。在本研究中,我们利用超高速离心成功分离了Maca-EVs,并对其形状、大小和标记物(CD63和TSG101)进行了详细分析。体内成像显示,Dil标记的Maca-EVs可以穿过脑血屏障并在脑内积累。行为学测试显示,Maca-EVs显著恢复了不可预测的慢性温和应激(UCMS)小鼠的抑郁样行为。UCMS小鼠粪便中g_Enterococcus、g_Lactobacillus和g_Escherichia_Shigella的丰度升高,这些菌属的丰度在服用Maca-EVs后显著恢复。Maca-EVs改变了UCMS小鼠粪便中代谢产物生物素、嘧啶和氨基酸(酪氨酸、丙氨酸、天冬氨酸和谷氨酸)的水平,这些代谢与五羟色胺(5-HT)的产生密切相关。Maca-EVs能够提高UCMS小鼠血清单胺类神经递质水平,其中5‐HT表现出最显著的变化。我们进一步证明,5‐HT通过调节GTP‐Cdc42/ERK通路,改善了脑源性神经营养因子BDNF的表达,并随后激活了TrkB/p‐AKT信号通路。这些结果表明Maca-EVs可能通过调节肠-脑轴来促进5‐HT的释放,从而改善抑郁行为。综上所述,我们的研究揭示了一种利用植物源细胞外囊泡治疗抑郁症的新方法。
视频解读
Bilibili:https://www.bilibili.com/video/BV1Hc411g79H/
Youtube:https://youtu.be/X0CrdB6qUy0
中文翻译、PPT、中/英文视频解读等扩展资料下载
请访问期刊官网:http://www.imeta.science/
全文解读
引 言
抑郁症是一种精神障碍疾病,影响着全球2.8亿人,其中包括5.0%的成年人和5.7%的老年人。严重的抑郁症会导致自杀,这是15-29岁人群死亡的第四大原因。世卫组织最近发布的数据表明,由于COVID-19的传播,全球抑郁症患病率增加了25%。研究发现,兴奋性突触、小胶质细胞、神经递质、神经免疫和神经炎症作用的功能障碍可能导致抑郁症的发展。因此,调节神经递质如血清素(5‐HT)、去甲肾上腺素(NE)、多巴胺(DA)等的药物被纳入抑郁症的临床治疗。传统的抗抑郁药,如单胺氧化酶抑制剂、选择性5-HT再摄取抑制剂、5-HT和去甲肾上腺素再摄取抑制剂已被证明具有特殊的抗抑郁功效。然而,其副作用如性功能障碍、恶心/呕吐、体重变化、睡眠中断和易上瘾等也不容忽视。此外,近50%的抑郁症患者对抗抑郁药物治疗没有疗效。因此,亟需开发安全有效的抗抑郁药。
在过去的十年里,越来越多的证据强调,肠-脑轴的作用在抑郁症的病理生理学发展中是必不可少的。这个轴指的是肠道和大脑之间的双向相互作用,由神经 (肠交感神经和迷走神经)、免疫(炎症细胞因子和细胞)和化学(微生物群代谢和神经递质) 信号促进。在抑郁症患者中,肠道和交感神经系统的活动升高。研究结表明,由脂多糖或膈下迷走神经切开术可减轻小鼠“抑郁症相关”微生物注射,证明了迷走神经在抑郁症中的潜在作用。此外,抑郁的特征是外周促炎细胞因子或趋化因子水平升高,如肿瘤坏死因子(TNF-α)、白细胞介素-6(IL-6)和IL-1β,激活的免疫细胞,如大脑中的神经元细胞和树突状细胞、肠道中的先天淋巴样细胞、体循环中的单核细胞和巨噬细胞。此外,某些肠道微生物群可以产生特定的代谢物(甲基色氨酸代谢,三甲胺-N-氧化物和短链脂肪酸),神经活性调节剂(5-HT,氨基丁酸,BDNF和神经胶质源性神经营养因子),以及肠细胞或细菌分解分泌的其他因子。因此,肠-脑轴正在成为治疗抑郁症的药物开发的新靶点。
Lepidium meyenii Walp,又名玛咖,在安第斯地区种植了至少2000年,自2002年以来成功引进中国,包括云南、新疆、吉林和西藏等省。玛咖是一种传统的食用药用植物,以其在激素平衡、调节性功能障碍和激活能量等方面的功效而闻名。玛咖含有丰富的必需营养素和生物活性成分,这些成分因其颜色和类型而异,从而具有多种生物功能。最常用和研究的三种类型是黄色玛咖、红色玛咖和黑色玛咖。最近,玛咖及其提取物因其潜在的神经保护和抗抑郁作用而引起了极大的关注。研究表明玛咖酰胺通过其抗炎、神经营养和突触保护功能修复皮质酮诱导的海马损伤。黄色、红色和黑色的玛咖在去卵巢的小鼠中也显示出了抗抑郁活性。此外,玛咖的石油醚提取物通过激活小鼠大脑中的去甲肾上腺素能和多巴胺能系统,抑制氧化应激,表现出抗抑郁作用。然而,玛咖及其提取物穿越血脑屏障(BBB)的有限能力显著阻碍了其作为治疗药物的潜力。植物来源的细胞外囊泡(PDEVs),类似于动物来源的细胞外囊泡(EVs),已被证明参与人类细胞间的通信,发挥多种生物学功能,如先天免疫和婴儿免疫信息调节。EVs的颗粒(< 200 nm)使它们能够自由地穿过血脑屏障,使它们有望成为转化应用的新候选物。在这项研究中,我们成功地从玛咖中分离和纯化了EVs(Maca‐EVs)。行为学实验表明,Maca‐EVs对不可预测的慢性轻度应激(UCMS)小鼠具有显著的抗抑郁作用。Maca-EVs的抗抑郁活性可能归因于血清5‐HT水平的提高和随后BDNF/TrkB/AKT轴的激活。血清5‐HT水平升高的可能机制包括肠-脑轴的调节等。
结 果
Maca‐EVs的分离与鉴定
如图1A所示,利用梯度超速离心法从新鲜玛咖汁中分离出Maca-EVs。透射电子显微镜成像和纳米颗粒分析显示,Maca-EVs是不规则的双层膜囊泡形状(图1B),平均直径为134 nm(图1C)。此外,与玛咖上清相比,Maca-EVs表达了更高的细胞外囊泡标记物CD63和TSG101(图1D),表明从玛咖果汁中成功分离了Maca-EVs。接下来,利用IVIS光谱成像系统研究了C57BL/6J小鼠体内对Maca-EVs的摄取。与未注射Maca-EVs的小鼠相比,静脉注射Dil标记的Maca-EVs(Maca-EVs-Dil)在小鼠大脑中积累了较强的红色荧光信号(图1E)。在小鼠接受Maca-EVs-Dil和free-Dil注射12h后处死小鼠并制备大脑冷冻切片。免疫荧光染色证实,Maca-EVs可以穿过血脑屏障并在大脑中积累(图1F)。

图1. Maca‐EVs的分离和表征
(A)Maca-EVs的分离流程图。(B)Maca-EVs透射电镜图像(红色箭头)。比例尺:200 nm。(C)NTA分析显示,Maca-EVs的尺寸约为134 nm。(D)Western blot分析在玛咖上清和Maca‐EVs中EVs标志物(CD63和TSG101)的表达。(E)IVIS光学成像系统对小鼠大脑中Free-Dil或Maca‐EVs-Dil在体内位置的代表性图像。(F)注射Free-Dil或Maca‐EVs-Dil后12 h死亡小鼠脑组织的代表性免疫荧光图像。标尺:100 μm。
Maca-EVs可逆转UCMS小鼠的抑郁样行为
接下来,我们在体内评估了Maca-EVs的抗抑郁样作用。我们在C57BL/6J小鼠中每日应用UCMS慢性温和刺激8周,建立抑郁模型(图2A)。UCMS体重显著降低(图S1A),在尾悬挂试验(图S1B)和强迫游泳试验(图S1C)中增加了静止时间,增加了小鼠梳理和进食潜伏期(图S1D,E),并减少蔗糖消耗(图S1F)。开放臂和闭合臂的时间没有明显变化,提示成功建立了抑郁模型,而不是焦虑模型(图S1G,H)。对照组小鼠随机分为两组:对照组和对照组注射Maca-EVs 100 μg/kg)。UCMS小鼠随机分为4组:UCMS 和UCMS注射不同剂量的Maca-EVs(50、100和200 μg/kg)。Maca-EVs注射剂量为:每2天给药1次,2周共8次。值得注意的是,在Maca-EVs治疗期间,每天持续给小鼠UCMS刺激。收集小鼠的粪便和血液样本,并在行为学测试完成后处死小鼠(图2A)。UCMS小鼠的体重持续下降,但与给予Maca-EVs(200 μg/kg)的UCMS小鼠体重与对照小鼠相当(图2B)。Maca-EVs可改善UCMS小鼠的抑郁样行为,如悬尾试验 (图2C)和强迫游泳试验(图2D)中不动时间延长。旷场试验(图2E)显示,Maca-EVs可以改善UCMS小鼠的探索行为,增加其行走距离(图2F)和进入中心区域的频率(图2G)。蔗糖消耗测试表明,与未处理的UCMS小鼠相比,Maca-EVs给药的UCMS小鼠的进食潜伏期(图2H)或梳毛潜伏期(图2I)显著减少,并且蔗糖偏好(图2J)增加。这些数据表明,Maca-EVs在治疗抑郁症方面具有潜在的治疗潜力。
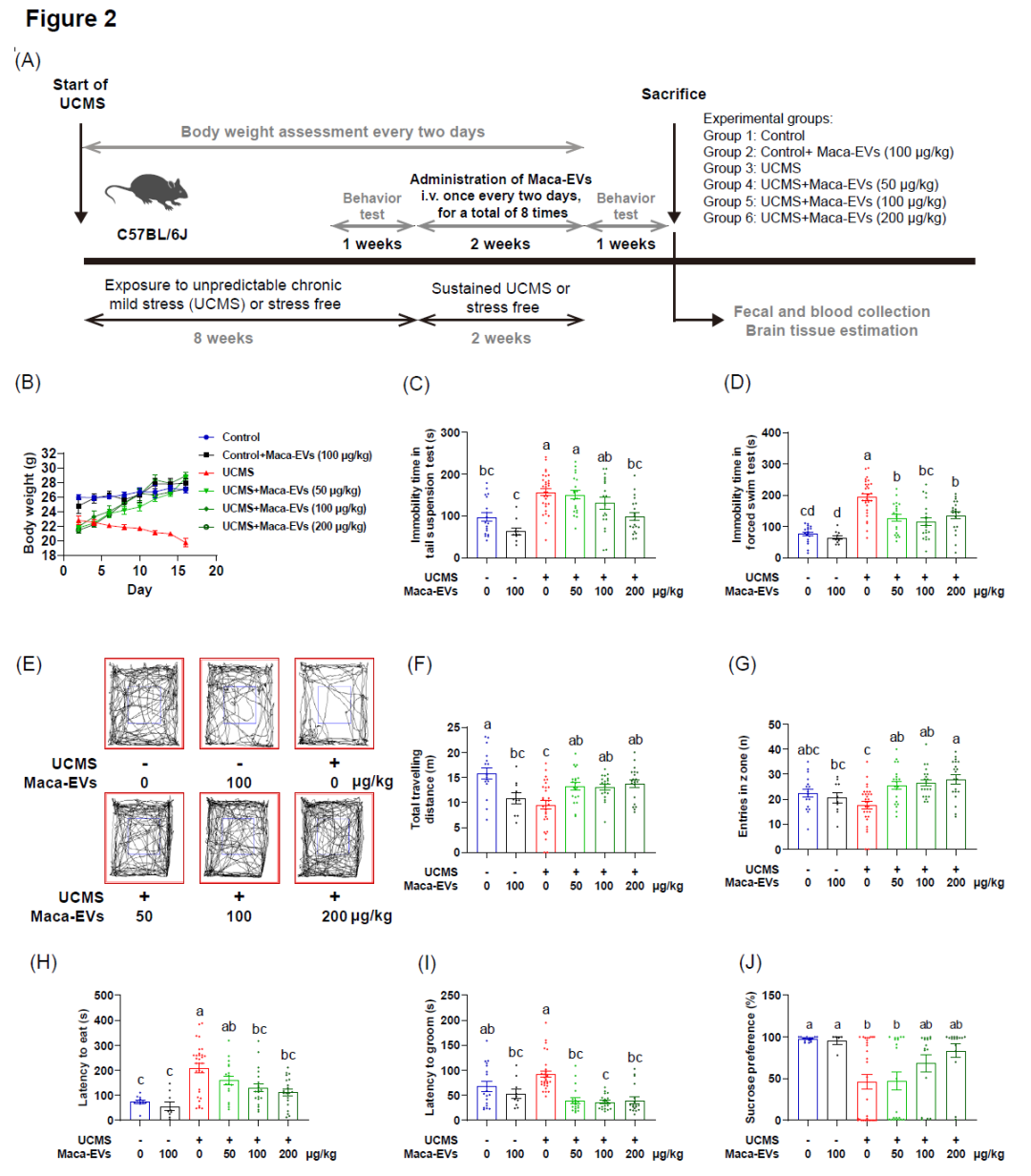
图2. Maca-EVs对UCMS小鼠的抗抑郁作用
(A)本研究中UCMS模型的建立和给药情况的示意图。(B)Maca-EVs处理或未处理的对照组和UCMS组小鼠治疗后在不同时间点的体重变化。对Maca-EVs处理或未处理的对照组和UCMS组小鼠进行行为学实验分析:小鼠悬尾试验(C)、强迫游泳试验(D)中静止时间、代表性开旷场照片(E)、露天试验总行进距离(F)、进入区域次数(G)、小鼠进食潜伏期(H)、梳理潜伏期(I)、蔗糖偏好百分比(J)。所有数据以均数±SEM表示(每组n=15-30个实验)。
Maca-EVs处理或未处理的对照组和UCMS小鼠粪便微生物群的变化
接下来,我们研究了四组小鼠(对照组、对照组+Maca-EVs、UCMS和UCMS+Maca-EVs)的肠道微生物群的动态变化。我们共获得了718,971条高质量的16S核糖体RNA(rRNA)序列,平均每个样本含有59,914个reads。我们将样本稀疏到相同的测序深度(每个样本56,402个reads)后进行聚类,共得到676,824个序列,将其分成了1434个操作分类群(OTU)进行下游分析。在门水平上,Bacteroidota和Firmicutes是所有类群中数量最多的(图3A),而在属水平上,Muribaculaceae是数量最多的(图3B)。乳杆菌属在不同组之间存在差异,与UCMS组相比,Control、Control+Maca-EVs和UCMS+Maca‐EVs组显著减少(p < 0.05,图3B)。相比之下,UCMS组的Akkermansia、Dubosiella和Faecalibaculum的丰度明显低于其他组(图3B)。这些数据表明,乳杆菌属、Akkermansia属、Dubosiella属和Faecalibaculum 属可能与UCMS和Maca‐EVs的作用有关。我们还研究了Maca-EVs对微生物多样性的影响,发现与对照组和UCMS组相比,Maca-EVs组(Control+Maca-EVs和UCMS+Maca-EVs)的微生物丰富度(Faith‐pd和Observed features index)更高(图3C,D)。只有Control+Maca-EVs组的Faith‐pd指数显著低于Control组(p < 0.05,图3D)。与对照组相比,UCMS组群落丰富度降低但不显著。为了评估不同组之间肠道微生物群β‐多样性的变化,我们进行了基于Bray‐Curtis距离矩阵的主坐标分析(图3E)和偏最小二乘‐判别分析(PLS‐DA)(图S2B)。通过ANOSIM检验(R = 0.772,p = 0.001),我们发现这四组被聚集成了不同的β多样性估计值组。为了更深入地了解Maca-EVs给药后微生物群的变化,我们使用线性判别分析效应大小(LEfSe)比较了16S重组DNA OTU水平上的微生物组成。Venn图(图S2A)显示,对照组和对照组+Maca‐EVs组共有444个OTU,对照组和对照组+Maca‐EVs组共有494个OTU,对照组和对照组+Maca‐EVs组共有222个OTU。对照组和UCMS组共有578个OTU,对照组共有360个OTU,UCMS组共有375个OTU。UCMS组和UCMS+Maca-EVs组共有471个OTU,UCMS组共有382个OTU,UCMS+Maca-EVs组共有209个OTU。我们将Control、Control+Maca-EVs和UCMS+Maca-EVs分为一个新组,称为New_Control。New_Control组与UCMS组之间共有11个差异OTU具有统计学意义(p < 0.05;线性判别分析> 4.0),如图3F所示,其中New_Control组富集1个OTU,UCMS组富集10个OTU。在这些差异OTU中,有2个OTU属于乳酸杆菌目,分别在科水平上属于乳酸杆菌科和肠球菌科。为了进一步研究与抑郁样症状相关的OTU,我们选择了New_Control组和UCMS组之间存在显著差异的四种典型细菌(图3G-J),观察它们在四组之间的丰度变化。此外,我们还选择了对照组和UCMS组之间存在显著差异的四种典型细菌(图S2C-F)。
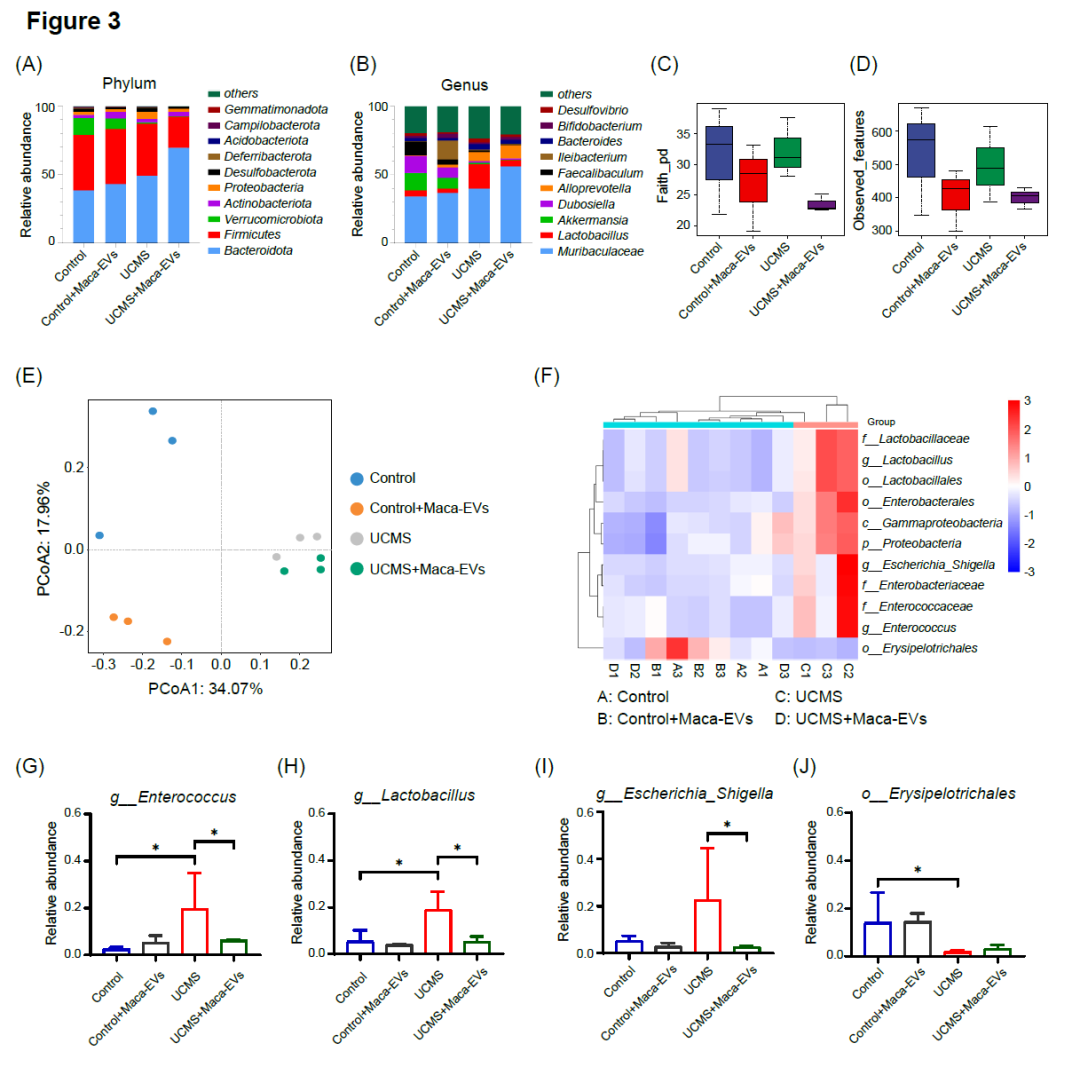
图3. Maca-EVs对照组和UCMS小鼠粪便中微生物群组成的影响
在行为测试完成后,我们处死小鼠并收集了粪便样本进行16S rRNA测序分析。本研究包括了对照组、对照组+Maca-EVs、UCMS组和UCMS+Maca-EVs组(每组n = 3个实验)。(A,B)展示了不同类群在门和属水平上的微生物群相对丰度(前10位)。(C,D)显示了Faith's系统发育多样性指数(pd)的Alpha多样性,并呈现其在不同类群中的观察特征。(E)展示了基于样本组间Bray Curtis距离矩阵的主坐标分析(PCoA)。通过采用线性判别分析效应大小(LEfSe),我们比较了各组之间在16S rDNA操作分类单位(OTU)水平上的微生物组成。结果显示,New_Control组与UCMS组之间有11个OTUs的差异具有统计学意义(p < 0.05;线性判别分析[LDA] > 4.0)。(G-J)显示了New_Control组与UCMS组之间典型细菌丰度的显著差异。
Maca-EVs处理或未处理的对照组和UCMS小鼠的粪便代谢特征的变化
鉴于肠道微生物组经常调节宿主的meta-Q代谢途径,在本研究中我们采用基于气相色谱-质谱(GC-MS)的代谢组学方法,对四个组别的代谢特征进行了比较。为了检测更多的代谢物,我们采用了非靶向代谢组学方法,包括正离子(pos)和负离子(neg)两种模式。通过主成分分析和偏最小二乘判别分析(PLS-DA),我们评估了不同组别粪便样本代谢特征的变化情况(见图S3A-D)。其中,图S3A和S3B为pos模式,图S3C和S3D为neg模式。总体而言,四个组别之间的代谢特征在空间上存在明显分离,尤其是New_Control组和UCMS组之间存在显著差异,表明它们在代谢物组成上存在差异。在pos模式下,我们鉴定出New_Control组与UCMS组之间的63种差异代谢物(VIP > 1,p < 0.05,|log FC| > 1)(见图4A),其中包括香草醛、L-5-羟色氨酸、4-(2,3-二氢-1,4-苯并二氧辛-6-基)丁酸、6-甲氧基-2-萘酸等;而UCMS组则富集了前列腺素I2、溶血酶22:5、N-乙酰氨酸、N-乙酰-天冬氨酸等代谢物。在neg模式下(见图S3G)。New_Control组与UCMS组之间存在47种差异代谢物(VIP > 1,p < 0.05,|log FC| > 1)。New_Control组富集的代谢物包括N-异丁基甘氨酸、N-乙酰丙氨酸、泛酸、2-酮-4甲基硫丁酸等,而UCMS组则富集了LPE 18:1、芹菜素、糖精、5-甲氧基水杨酸等代谢物。我们选择了四种典型代谢物(香兰素、L-5-羟色氨酸[5-HTP]、4-(2,3-二氢-1,4苯并二氧辛-6-基)丁酸和6-甲氧基-2-萘酸),发现它们在New_Control组和UCMS组之间存在显著差异,并分析了它们在四个组别之间的变化情况(见图4B-E)。途径富集分析结果显示,UCMS导致了与生物素、嘧啶、酪氨酸、丙氨酸、天冬氨酸和谷氨酸代谢相关的途径发生显著改变(见图S3E)。通过色氨酸羟化酶将色氨酸转化为5-羟色氨酸(5-HTP),然后通过5-HTP脱羧酶进一步转化为5-HT(见图S3F)。在我们的代谢组学分析中,我们发现UCMS组中5-HTP的含量显著降低,而UCMS + Maca-EVs组中5-HTP的含量较高(见图4C),这表明UCMS可能影响5-HT的生物合成。综上所述,这些结果表明代谢物与不同组别之间存在关联。为了研究改变的肠道细菌和代谢物之间潜在的相互作用,我们基于Spearman相关分析构建了一个共发生网络(见图4F)。在pos模式下,我们发现乳酸杆菌属与粪便代谢物之间存在强烈的共发生关系,其中代谢物包括2-胡椒二苯甲酸、R-1甲烷酰磷酸、PPK、异香草酸和N6-乙酰-L-赖氨酸。肠球菌属与粪便代谢物之间也存在密切的共发生关系,其中代谢物包括LPC15:0、D-(--)-水杨苷、ACar16:1、4-孕烯-17,20-二醇3-1、LNK和N6-乙酰-L-赖氨酸。志贺氏杆菌属与粪便代谢物孕烯醇酮、三角三内酯、雌二醇、2-脱氧尿苷和脱氧胞苷之间形成了密切的共生关系。乳酸杆菌属与PPK、N6-乙酰-L-赖氨酸呈正相关,与2-胡椒二苯甲酸、R-1磷酸甲胺和酰苯甲酸呈负相关。肠球菌属与4-Pregnen-17、20-Diol-3-One、LNK和N6-acetyl-L-赖氨酸呈正相关,与LPC 15:0、D-Salicin和ACar 16:1呈负相关。志贺氏杆菌属与2-脱氧尿嘧啶和脱氧胞苷呈正相关,与孕酮、三角三内酯和雌酯酸呈负相关。综上所述,我们的研究结果表明改变的肠球菌、乳酸杆菌和志贺氏杆菌与(pos)模式下的生物素和嘧啶代谢相关。在(neg)模式下,我们同样观察到共发生网络的存在,这表明改变的肠球菌、乳酸杆菌和志贺氏杆菌与酪氨酸、丙氨酸、天冬氨酸和谷氨酸代谢相关(见图S3F,G)。需要注意的是,这些发现揭示了改变的肠道微生物群与代谢物在New_Control组和UCMS组之间形成了协同和节点相关的共发生网络。
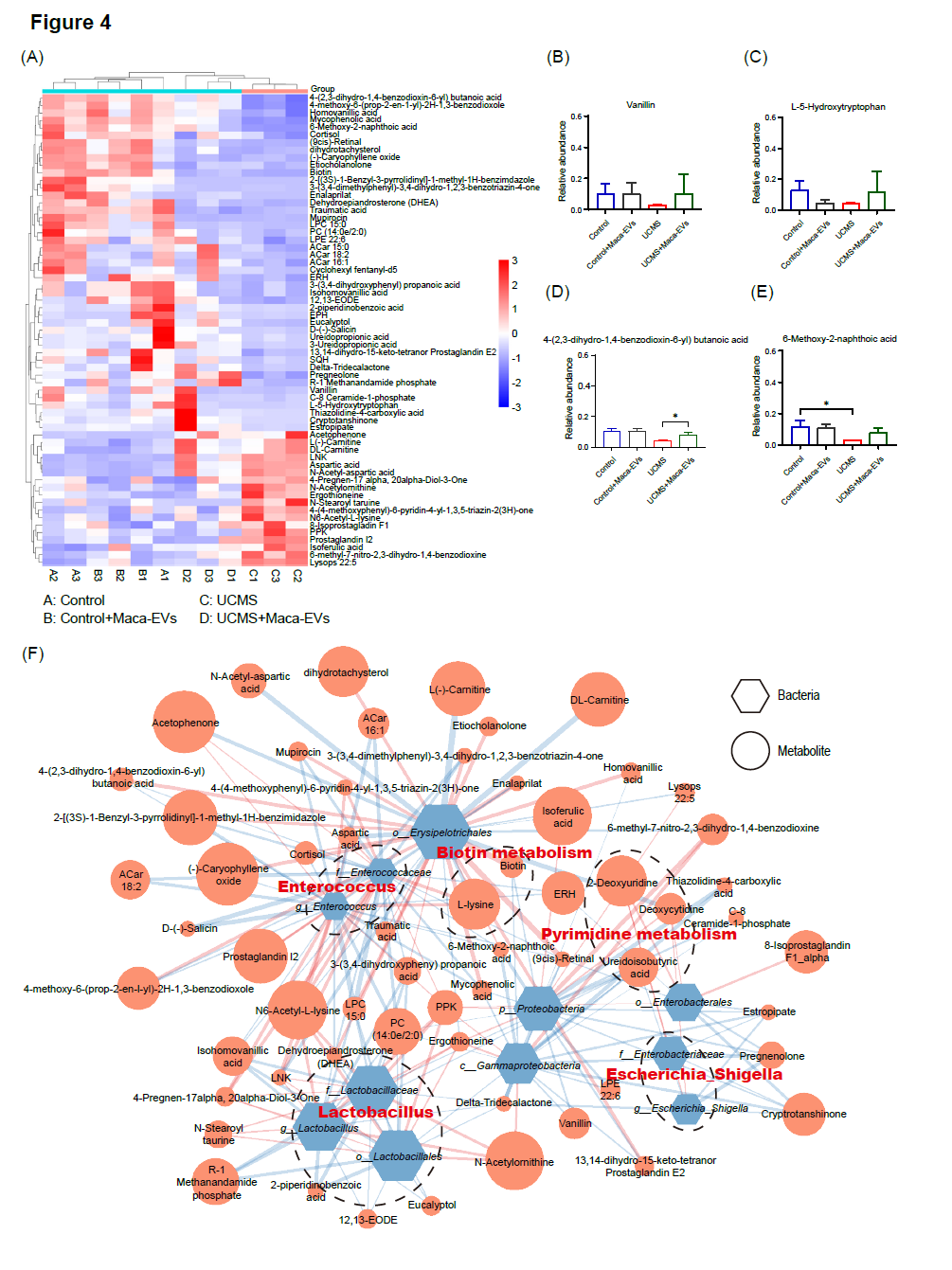
图4. Maca-EVs对对照组和UCMS小鼠粪便代谢组学的影响
(A)在pos模式下,我们发现New_Control组与UCMS组在代谢物水平上存在差异(VIP > 1,p < 0.05,|log FC| > 1)。(B-E)样品组间比较显示New_Control组与UCMS组在4种典型代谢物水平上存在显著差异。(F)通过基于Spearman相关分析的共发生网络,我们确定了肠道细菌与血清代谢物之间的相互作用关系。
Maca-EVs可增强UCMS小鼠血清5-HT的表达
由于鉴定出的代谢改变与5-HT的产生密切相关,我们下一步研究了Maca-EVs是否调节单胺神经递质的产生。我们在完成行为测试后收集小鼠血清,通过高效液相色谱法测量三种主要单胺类神经递质(5-HT,NE和DA)的水平。NE和DA的信号太低,无法量化。我们观察到与对照组小鼠相比,UCMS小鼠血清5-HT水平显著下调,证实了UCMS模型的成功建立(图5A,B)。在三种剂量下,Maca-EVs均显著增加了UCMS模型的血清5-HT水平。与对照组相比,UCMS小鼠血清炎症因子IL‐1β、IL-6和TNF‐α水平升高,但差异不显著(图5C-E)。在200 μg/kg剂量下,Maca-EVs显著降低血清中TNF‐α水平,而IL‐1β和IL-6表达的降低无统计学意义。总的来说,血清单胺神经递质水平,特别是5-HT而不是炎症因子,可能有助于Maca-EVs的抗抑郁作用。

图5. 血清单胺类神经递质和炎症因子表达分析
(A)单胺因子的UPLC-MS/MS色谱图,包括标准品和对照小鼠血清中的5‐HT,以及经Maca-EVs治疗的UCMS小鼠血清。(B)经Maca-EVs治疗的对照组和UCMS小鼠血清中5‐HT浓度的定量数据。炎症因子浓度的定量数据,包括IL‐1β (C)、IL‐6(D)和TNF‐α(E)在对照组和经Maca-EVs治疗的UCMS小鼠血清中。所有数据均以Mean±SEM表示(每组n = 3-11个实验)。通过普通的单因素方差分析(ANOVA)评估显著性,然后在(B,C,E)中进行Tukey多重比较检验,在(D)中进行Kruskal-Wallis检验和Dunn多重比较检验。用不同字母表示的平均值表示组间显著性差异(p < 0.05)。
Maca-EVs通过激活UCMS小鼠海马和皮质中的GTP-Cdc42/ERK信号通路来减弱BDNF表达的下降
5-HT和BDNF信号的协同调节已经得到证实,其中5-HT激活BDNF表达,5-HT对神经元的生长和存活至关重要。异常的5-HT和BDNF信号与抑郁症的进展有关。给药剂量为200 μg/kg的Maca-EVs显著增加了血清5-HT水平,促使我们研究其是否会增强UCMS小鼠BDNF的表达。通过免疫荧光染色和WB分析,我们证实了UCMS小鼠海马(图6A,B)和皮质(图S4A,B)中BDNF表达的下调,为了进一步探索其潜在机制,我们研究了调节BDNF表达的GTP-Cdc42/ERK信号通路。WB分析显示,与注射生理盐水小鼠相比,服用Maca-EVs的UCMS小鼠海马(图6C)和皮质(图S4C)中GTP-Cdc42/Cdc42和p-ERK/ERK的比例上调。这些结果表明,Maca-EVs通过触发UCMS小鼠海马和皮质中的GTP-Cdc42/ERK信号通路来减弱BDNF表达的下降。
Maca-EVs通过调节UCMS小鼠海马和皮质的TrkB/AKT信号通路来减轻神经元数量的下降
此外,我们还探讨了BDNF表达上调是否会调节UCMS小鼠的神经元生长。BDNF与其受体酪氨酸激酶受体B(TrkB)相互作用并启动下游AKT信号传导以维持神经元生长。我们在UCMS小鼠中观察到TrkB表达下调和p-AKT/AKT比值上调,Maca-EVs显著升高TrkB水平而降低p-AKT/AKT比值(图6D和S4D,E)。免疫荧光染色显示,Maca-EVs给药可促进UCMS小鼠齿状回区神经元生长(图6E和S4F)。因此,数据表明,Maca-EVs通过调节皮层中的TrkB/p-AKT信号通路减轻神经元数量的减少。
5‐HT在体外通过调节GTP‐Cdc42/ ERK通路诱导BDNF表达并随后激活TrkB/AKT信号通路
为了确定5-HT激活是否调节GTP-Cdc42的活性,我们用ML141(GTP-Cdc42抑制剂)处理PC12细胞,观察到GTP-Cdc42/Cdc42和p-ERK/ERK的比值显著降低(图6F)。我们还发现,ML141处理可减弱5‐HT诱导的BDNF表达上调(图6G,H),这表明5‐HT通过调节GTP‐Cdc42/ERK通路刺激BDNF表达。此外,ML141处理阻断了5‐HT随后的TrkB/AKT信号传导,TrkB表达和p‐AKT/AKT比值下降证明了这一点(图6I,J)。这些数据表明,5‐HT激活GTP‐Cdc42/ERK通路,刺激BDNF表达并随后激活TrkB/AKT信号。
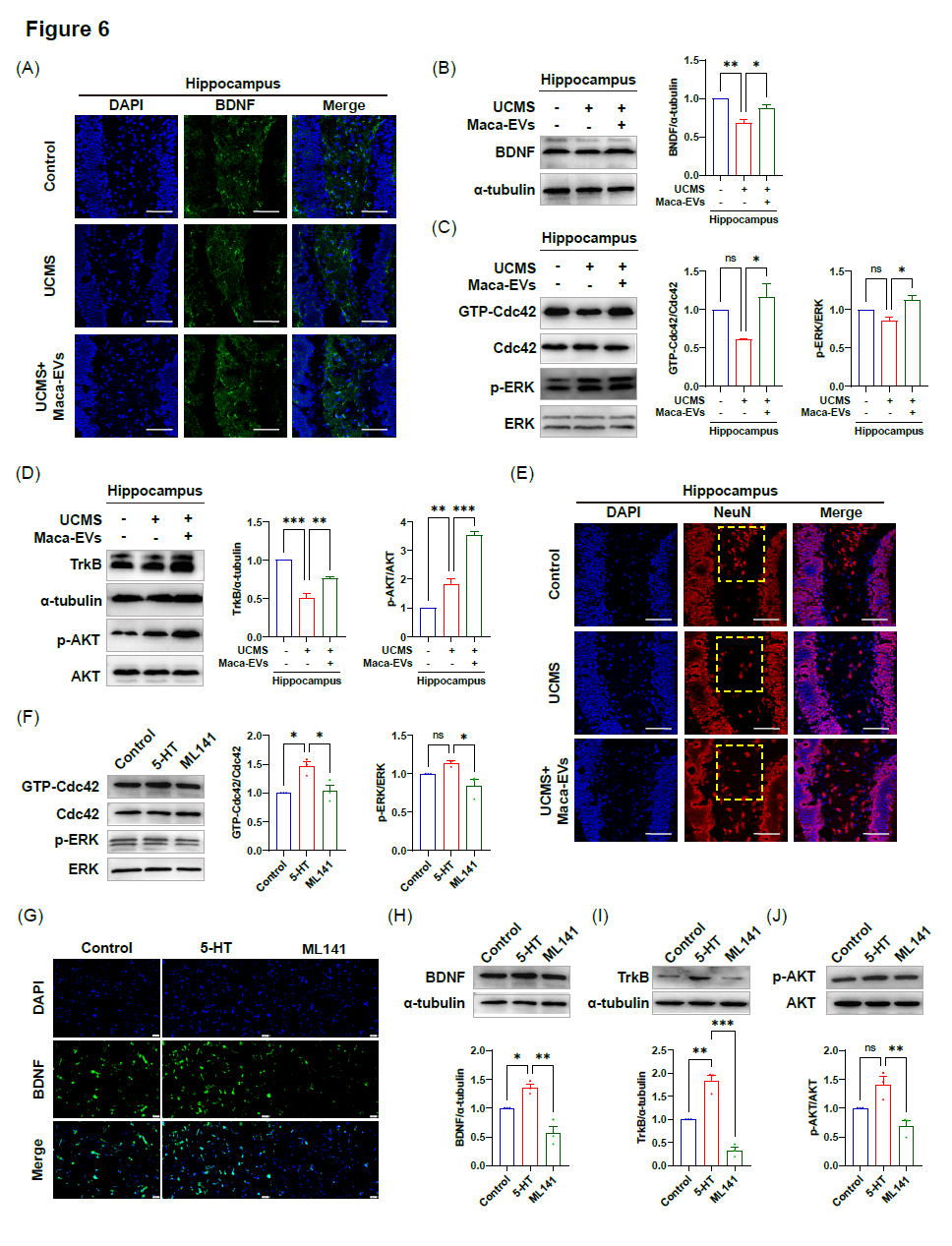
图6. Maca-EVs通过5‐HT调节小鼠海马BDNF/GTP Cdc42/ERK和TrkB/AKT信号通路
(A)对照组、UCMS组、Maca‐EVs(200 μg/kg)+UCMS组小鼠海马中BDNF表达的代表性免疫荧光图像。标尺:50 μm。对照组、UCMS组、Maca‐EVs(200 μg/kg)+UCMS组小鼠海马中BDNF(B)、Cdc42酶活性、p‐ERK和ERK(C)、TrkB、p‐AKT和AKT(D)表达的Western blot图像和定量数据。(E)对照组、UCMS组、Maca‐EVs(200 μg/kg)+UCMS组小鼠海马NeuN细胞的代表性免疫荧光图像。黄色虚线框为海马齿状回区。标尺:50 μm。(F)用5‐HT或ML141(一种GTP‐Cdc42抑制剂)处理PC12细胞12小时,再培养24小时。显示PC12细胞中GTP‐Cdc42、Cdc42、p‐ERK和ERK表达的Western blot图像和定量数据。免疫荧(G)和western blot分析(H)及BDNF在PC12细胞中的表达。标尺:50 μm。PC12细胞中TrkB (I)、p-AKT和AKT(J)表达的Western blot图像和定量数据。所有数据均以Mean±SEM表示(每组n = 3个实验)。在(B,C,D,F,H,I,J)采用单因素方差分析(ANOVA)和Tukey多重比较检验评价两组间的显著性,p > 0.05,*p < 0.05,**p < 0.01,***p < 0.001。
讨 论
据我们所知,我们是第一个成功地从玛咖中分离和表征Maca‐EVs。Maca‐EVs表现出与动物细胞源性EVs相似的特征,可以穿过血脑屏障。静脉注射Maca‐EVs显著降低了UCMS小鼠的抑郁行为。我们使用16S rRNA和基于GC-MS的代谢组学分析发现,Maca‐EVs改变了UCMS小鼠与抑郁症相关的粪便微生物群的组成,并改善了与5-HT产生相关的代谢,表明Maca‐EVs具有调节肠脑轴的潜力。此外,通过HPLC分析,发现UCMS小鼠血清5‐HT水平降低,而Maca‐EVs显著提高了UCMS小鼠血清5‐HT水平。体内和体外实验表明,5‐HT刺激BDNF的表达,进而激活BDNF/TrkB/AKT信号,这可能有助于Maca‐EVs的抗抑郁作用。
传统中医长期以来被广泛应用于抑郁症的治疗,特别是在亚洲。中药作用的分子机制涉及调节单胺传递、下丘脑-垂体-肾上腺轴、新营养因子(如BDNF)以及突触的数量和功能。据报道,玛咖提取物因其抗氧化活性而具有神经保护和抗抑郁作用。后来的研究发现,玛酰胺是通过抑制脂肪酸酰胺水解酶(脂肪酸酰胺水解酶调节神经祖细胞增殖)起到神经保护作用的活性成分。此外,研究表明玛咖提取物通过降低玛咖使用者体内的炎症细胞因子IL-6而具有抗炎特性。然而,玛咖及其提取物治疗抑郁症的确切分子机制仍有待阐明。在过去的几十年里,PDEVs是一种球形、椭圆形或杯状的囊泡,含有生物活性成分,包括脂质、蛋白质、核酸和次级代谢物,在治疗药物开发中获得了大量关注。越来越多的体外和体内研究表明,PDEVs具有抗癌、抗炎、抗氧化和再生活性。值得注意的是,Teng等人发现,来自生姜衍生PDEVs的microRNA(miRNA)可以靶向肠道细菌鼠李糖乳杆菌基因组,产生更多的吲哚-3-甲醛,从而促进IL-22的分泌,增强肠道屏障功能。尽管临床前研究前景看好,但PDEVs从实验室到临床的转化仍处于起步阶段。目前,美国路易斯维尔大学已经使用四种PDEVs(姜黄素、生姜、芦荟、葡萄)进行了三项临床试验(NCT04879810、NCT01668849、NCT01294072)。无论如何,这些临床试验的完整结果尚未被报道。因此,需要在PDEVs的分离纯化、理化特性、质量控制、安全性和递送途径等方面进行大量的研究。我们的团队之前已经从瓜科植物的果实苦瓜中分离EVs,并证明了其在治疗大脑脑缺血/再灌注和辐射诱发的心脏病方面的治疗潜力。因此,我们旨在从黑玛咖中分离出EVs,并在UCMS小鼠模型中研究其抗抑郁能力,以及相关机制。我们通过梯度离心成功分离了Maca-EVs,其表现出与动物细胞源性EVs相似的特征(图1A-D)。使用广泛用于标记EVs的亲脂性荧光染料Dil来表征和监测Mca‐EVs的细胞摄取或组织分布。体内成像显示,Dil‐Maca‐EVs可有效穿过血脑屏障,并在小鼠脑内具有更高的荧光密度(图1E,F)。此外,除了对蔗糖偏好的调节外,我们还通过行为测试(如悬尾测试、强迫游泳测试、开旷场地测试和蔗糖消耗测试)证明了Maca‐EVs对抑郁症UCMS小鼠的治疗效果(图2)。
肠脑轴代表中枢神经和消化系统之间的双向通信,涉及神经递质、神经免疫、神经内分泌和感觉神经通路。大量数据支持肠脑轴连接在抑郁症疾病环境中的作用。在微生物群耗尽的大鼠中,移植研究抑郁症患者粪便微生物群诱导的抑郁样行为和生理特征,提示肠道微生物群在抑郁症发展中的因果作用。此外,越来越多的努力已经致力于描绘抑郁症患者的微生物群组成和代谢的特征。Raes等人发现产生丁酸盐的Faecalibacterium和Coprococcus细菌在抑郁症患者中不存在。在这项研究中,我们发现乳酸杆菌属在UCMS小鼠中显示出更高的丰度,而Akkermansia,Dubosiella和Faecalibaculum在对照小鼠中的丰度高于UCMS小鼠,这表明它们在Maca‐EVs的抗抑郁作用中可能发挥作用(图3A,B)。此外,四组微生物群的α‐多样性和β‐多样性也有所不同(图3C-E)。通过将Control、Control+ Maca‐EVs和UCMS+Maca‐EVs重组为一个新的组(New_Control),我们进一步证明,Maca‐EVs降低了g_Enterococcus和g_Lactobacillus的丰度,而在UCMS小鼠中,这两种细菌的丰度增加(图3G,H)。大多数人类肠道菌群分析发现,抑郁症患者中g_Enterococcus和g_Lactobacillus的丰度减少。然而,它们在抑郁小鼠中的丰度并没有达成一致。在Bharwani和研究中发现g_Enterococcus和g_Lactobacillus的丰度增加,而在Farshim等和Galley等的研究中发现丰度降低。必须承认,由于个体和物种的差异,肠道微生物群组成的丰度、α‐多样性和β‐多样性存在不一致。Yang等人描述了在重度抑郁症患者中,g_Bacteroides显著升高,g_Blautia和g_Eubacterium显著下降,同时γ-氨基丁酸盐、苯丙氨酸和色氨酸代谢发生改变。这里,Pathway富集分析表明,UCMS代谢途径的主要改变与生物素、嘧啶、酪氨酸、丙氨酸、天冬氨酸和谷氨酸代谢有关(图4F和S3E)。酪氨酸代谢相关产物L‐5‐羟色氨酸水平在UCMS小鼠中下降,但在给药Maca‐EVs后升高(图4C)。这表明Maca‐EVs通过调节肠道菌群组成和代谢来提高血清5‐HT水平。兴奋性或抑制性神经递质在神经元兴奋性的稳态调节中起重要作用,赋予正常的脑功能。兴奋性神经递质5-HT水平或活性降低与抑郁症相关的理论仍然具有影响力。最近,Moncrieff等人暗示,5‐HT水平或活动的降低可能不是抑郁症的原因。然而,目前治疗抑郁症的处方药主要是通过提高使全身兴奋性神经递质5‐HT、NE和DA水平起作用。Rizzo等人证明,选择性5-羟色胺再摄取抑制剂氟西汀可增强5-羟色胺的表达,促进神经干细胞的增殖,而5-羟色胺受体拮抗剂可逆转这一过程。在我们的研究中,所有剂量的Maca‐EVs均显著提高了UCMS小鼠血清5‐HT水平(图5B)。先天和适应性免疫功能障碍已被证明与抑郁症的病理生理有关。Grosse等发现单核细胞基因表达在重度抑郁症患者中与年龄相关。因此,通过检测三种特异性血清炎症细胞因子IL‐1β、IL‐6和TNF‐α来研究Maca‐EVs是否会调节炎症反应,这三种特异性血清炎症细胞因子已被证明与抑郁症呈正相关。在我们的模型中,在UCMS小鼠中检测到血清IL-1β、IL-6和TNF-α升高,但没有统计学意义(图5C-E)。这可能是因为在抑郁诱导的后期阶段测试了炎症因子。然而,Maca‐EVs仅在200 μg/kg剂量下显著降低IL-6和TNF-α水平。这些数据表明,5‐HT信号对Maca‐Evs治疗更为敏感,接下来将重点研究其在治疗抑郁症UCMS小鼠中的作用。
5‐HT和BDNF在调节突触可塑性方面存在协同作用,5‐HT触发BDNF的表达,BDNF促进5‐HT神经正常生长和存活,维持成人脑突触可塑性。5-HT信号的激活需要与5-HT受体相互作用,该受体不仅偶联而且激活G-蛋白(Rho,Rac和Cdc42)相关途径。Udo等人证实5-HT激活了应用感觉神经元中的GTP-Cdc42,促进了学习相关突触的生长。Elena等人发现,在神经母细胞瘤细胞中,5-HT受体的激活通过Cdc42介导的途径导致丝状足形成,并伴有RhoA相关的细胞围成。Errico等和Speranza等研究表明,5‐HT信号激活可通过ERK通路增强神经元功能。其他人也发现(R)‐氯胺酮异构体通过激活ERK‐NRBP1‐CREB‐BDNF通路发挥抗抑郁作用。事实上,在注射了Maca‐EVs的UCMS小鼠的皮层和海马中,GTP‐Cdc42/Cdc42和p‐ERK/ERK以及BDNF的表达比例升高(图6A-C和S4A-C)。有证据表明,BDNF信号通过其受体TrkB是抗抑郁药物的作用所必需的。Adachi等人发现TrkB缺失而BDNF缺失会导致抑郁症治疗能力下降。He等人的研究表明,Alpinia oxyphylla Miq激活BDNF/TrkB/AKT信号通路可促进海马神经发生。因此,还发现Maca‐EVs增加了UCMS小鼠皮层和海马中TrkB和磷酸化AKT的表达(图6D和S4D,E)。体外实验进一步证实,5-HT通过GTP Cdc42/ERK通路增强BDNF表达,进而刺激TrkB/AKT信号通路(图6F-J)。
遗憾的是,我们只在最初分离了Maca‐EVs并研究了其抗抑郁作用。观察到Maca‐Evs可以影响粪便微生物群组成,提高血清5‐HT水平,通过激活GTP‐Cdc42/ERK信号通路,导致BDNF表达增加。肠黏膜中的肠嗜铬细胞是5‐HT合成、储存和释放的主要部位。揭示了Maca‐EVs在调节粪便微生物群/代谢物产生5‐HT中的潜在作用,这需要进一步的实验来阐明。然而,Maca‐EVs可以穿过血脑屏障,它们可能通过直接影响大脑或通过外周途径发挥抗抑郁治疗作用。在这项研究中,我们只关注了Maca‐EVs对肠脑轴调节的影响。未来的研究将采用灌胃或靶向给药的方式给药Maca‐EVs,以阐明Maca‐EVs治疗抑郁症的潜在机制。在本研究中还有其他几个局限性。首先,Maca‐EVs产生类似于玛咖或其提取物的抗抑郁作用,可能是因为Maca‐EVs含有它们。值得注意的是,Maca‐EVs的纳米级尺寸(< 200nm)使它们能够克服玛咖提取物成分的缺点,如多糖、蛋白质和生物碱(这些成分是高分子和不溶性的),从而使它们能够穿过血脑屏障或肠道屏障。然而,应该进行彻底的组学研究,以了解生物活性成分,如核酸、蛋白质、脂质和次级代谢物,以验证这一推测。此外,赋予Maca‐EVs抗抑郁机制的关键因素仍未被探索。最近,含有miRNA的EVs作为下一代药物在商业领域引起了极大的兴趣。然而,单个miRNA可能不如多个miRNA有效。即使一些miRNA显示出良好的治疗能力,但由于其易于降解,多靶点,以及潜在的生物安全风险,其开发成为药物具有挑战性。例如,评估MRX34(miRNA‐34)治疗黑色素瘤疗效的1期临床试验(NCT02862145)由于严重的免疫相关副作用而被撤销。新兴研究指出,Evs的治疗活性源于其内容物的协同作用。因此,确定在Evs功能中起重要作用的精确元素似乎不那么重要。其次,我们只揭示了5‐HT/BDNF在Maca‐EVs抗抑郁活性中的作用。除BDNF外,其他神经营养因子包括血管内皮生长因子、成纤维细胞生长因子2、胰岛素样生长因子1和激活素A也值得研究。其他两种单胺类神经递质在抗氧化或抗炎作用中的作用机制需要进一步研究。最后,我们进行了一项可能影响肠道菌群组成的蔗糖测试来评估抑郁行为,尽管我们的数据显示Maca‐EVs并不影响蔗糖偏好水平。在未来的研究中,我们将避免这些技术问题,以研究Maca‐EVs调节肠-谷物轴的确切机制及其与Maca‐EVs治疗抑郁症的相关性。
方 法
动物
雄性C57BL/6J小鼠,体重18-22 g,购自北京维通利华实验动物科技有限公司。本研究经徐州医科大学机构动物保护委员会(202006W037)批准。所有动物实验均按照机构和国家指导方针进行。
Maca-EVs 的分离
用去离子水仔细洗涤三次,挤压成汁。在4℃条件下,1000×g离心10 min, 3000×g离心20 min,10000×g离心40 min。然后将上清液以150000×g超离心90分钟(Optima XE-90,Beckman Coulter Life Sciences,Indianapolis,美国)。将沉淀重悬于磷酸盐缓冲盐水(PBS)中,然后转移到梯度蔗糖溶液(8 %、30 %、45 %和60 %)中,在150000×g下超离心90分钟。收集30 %至45 %蔗糖层之间的条带,用PBS洗涤。然后在150000×g超离心90min,用0.22 um滤膜(#SLGV004SL,Millipore)通过,得到无菌Maca-EVs。使用BCA检测试剂盒(#P001,Beyotime)对Maca-EVs进行蛋白定量。
透射电子显微镜
将Maca-EVs(10 μl)吸附在碳包覆网格上1 min,然后用一滴水洗涤,用1 %醋酸铀酰染色15 s。利用透射电镜(FEI Tecnai G2 Spirit Twin,New York,USA)检测吸收的Maca-EVs。
纳米颗粒跟踪分析
利用纳米颗粒跟踪分析仪(ZetaView,Bavaria,Germany)检测了Maca-EVs的大小分布。根据说明书,用PBS稀释Maca-EVs,然后加入分析细胞开始测定。得到了尺寸分布的数据。
Maca-EVs的体内摄取
将分离的Maca-EVs与亲脂荧光染料Dil(Molecular Probes,Oregon,USA)室温孵育30 min。将Dil标记的Maca-EVs(Maca-EVs-Dil)在150 000×g下超离心90 min,丢弃上清,用200 μl PBS重悬。C57BL/6J小鼠静脉注射Maca-EVs-Dil或游离Dil(200 μg/kg),于12 h通过IVIS®光谱系统(Perkin Elmer,Massachusetts,USA)成像。
不可预测的慢性轻度应激(UCMS)模型和给药方案
根据文献报道建立UCMS模型和给药设计。根据报道,每天随机给予C57BL/6J小鼠2或3种轻度不可预测的应激源,持续8周,建立UCMS模型。有12种应激源,包括夹尾5分钟、0°C冷泳5分钟、45°C笼12小时、约束6小时、剥夺食物或水12小时、潮湿笼24小时、空笼12小时、高速振动4小时、闪光灯照射12小时、笼内垫料污染6小时、昼夜光照循环反转24小时、猫叫声、毛发4小时。每个压力源不是连续施加的。对照组和UCMS小鼠分别饲养。每2天称一次体重,直至处死。
随后,每2天静脉注射1次Maca-EVs(0、50、100、200 μg/kg),连续2周。重要的是,在两周的Maca-EVs治疗期间,每天维持UCMS模型,由于小鼠的高度自我修复能力,避免了UCMS的潜在恢复。以注射Maca-EVs(0、100 μg/kg)的无应激小鼠为对照。最后,在行为测试完成后,处死小鼠,获得粪便颗粒、血液样本和脑组织。每只小鼠单独饲养在消毒盒中收集新鲜粪便颗粒。200 mg的粪便样品在-80°C快速冷冻保存直至处理。将血样在4℃下3500 rpm离心10-15 min,收集血清。然后将血清转移到无菌EP管中,在-80°C保存以供进一步分析。新鲜脑组织用4 %多聚甲醛在4°C下固定过夜。然后将其浸泡在15 %和30 %蔗糖溶液中完全脱水,包埋在OCT化合物(#4583 Tissue-Tek®O.C.T. compound,Sakura)中,制备冷冻切片(14 μm)。新鲜脑组织也保存在冰冷的盐水溶液中。将海马和皮质组织分离于冷盘上,立即液氮冷冻,-80°C保存,用于western blot检测。海马和皮质也分别用含有1%磷酸酶和蛋白酶抑制剂(#78442,Thermo Scientific)的冷冻RIPA缓冲液(#89900,Thermo Scientific)匀桨。然后在16000×g 4℃下离心15 min,得到的上清液用于测定。
悬尾实验
将小鼠倒挂,尾巴用胶带固定在支架上,置于ST-100尾悬室(Tai-Co软件,中国成都)中。适应1 min后,使用Tail Suspension Scan TM(Clever Sys Inc.,VA,USA)记录并分析小鼠在随后5 min(活动阈值=30%)内的静止时间。
强迫游泳实验
将小鼠分别置于一个玻璃筒(直径10 cm,Tai League Software,成都,中国)中,该玻璃筒中装满水(25±1°C),深度为14 cm,放置6分钟。使用Tail Suspension Scan TM(Clever Sys Inc.,VA, USA)记录并分析最后5分钟的静止时间。当小鼠放弃逃跑并保持头露出水面时,被认为是不动的。
开旷场实验
将每只小鼠小心地置于62.5 × 74 × 51 cm的开场室中央,用摄像机记录5 min的行为。使用OFT软件(SMART 3.0,Panla,Spain)对总行驶距离和进入区域的数据进行分析。
新奇喂食实验
NSFT按照前面的描述进行。将小鼠置于50 cm×50 cm×20 cm的白色塑料箱中,箱底铺上木质被褥(每只小鼠换一次),禁食24 h。将单个食物颗粒(普通食物)放在一张直径为10厘米的滤纸上,放置在明亮照明(~500勒克斯)的容器中心。老鼠被放置在盒子的一个角落,在10分钟内测量进食的潜伏期。进食的定义是咬,而不是简单地嗅或触摸食物。试验结束后,这些动物立即被转移到它们自己的笼子里。
喷溅实验
泼溅试验是将10 %的蔗糖溶液喷洒在家笼小鼠的背毛上。蔗糖溶液会弄脏皮毛,增加梳理的时间。记录小鼠梳理的时间为6分钟。
糖水偏好实验
在24小时的断水后,每只小鼠暴露在两瓶中12小时,一瓶含有去离子水,另一瓶含有1 %的蔗糖溶液。两个瓶子的位置随机放置,并在6小时后互换,以避免可能的侧边偏好。蔗糖偏好百分比由蔗糖溶液消耗量与总液体消耗量之比计算。
高架十字迷宫实验
EPM测试用于评估啮齿动物的焦虑相关行为。EPM装置包括位于地板上方的“+”形模组,具有两个张开臂(OA)和两个闭合臂(CA),通过中央平台连接。将小鼠置于迷宫中央,通过安装在迷宫上方的摄像机记录小鼠的行为,并使用视频跟踪系统对其进行分析。对张开双臂或紧闭双臂的时间进行评分,以衡量焦虑类行为。
16S rRNA测序
采用CTAB/SDS法提取粪球总基因组DNA。在1 %琼脂糖凝胶上监测DNA浓度和纯度。根据浓度,用无菌水将DNA稀释至1 ng/µl。利用条形码特异引物(如16S V4: 515f - 806r、18S V4: 528F-706R、18S V4: 1380F-1510R等)扩增不同区(16S V4/16S V3/16S V3-V4/16S V4-v5、18S V4/18S V9、ITS1/ITS2、Arc V4)的16S rRNA/18SrRNA/ITS基因。所有PCR反应用15 µl Phusion®High-Fidelity PCR Master Mix(New England Biolabs), 0.2 µM正向和反向引物,约10 ng模板DNA进行。热循环包括98°C初始变性1 min,然后是98°C变性10 s, 50°C退火30 s, 72°C延伸30 s,最后72°C延伸5 min的30个循环。将相同体积的1X上样缓冲液(含SYB绿色)与PCR产物混合,在2%琼脂糖凝胶上进行电泳检测。PCR产物以等密度比混合。然后用Qiagen凝胶提取试剂盒(Qiagen,Germany)纯化混合PCR产物。使用TruSeq®DNA PCR-Free Sample Preparation Kit(Illumina, USA)按照制造商的建议生成测序文库,并添加索引代码。使用Qubit@ 2.0荧光仪(Thermo Scientific)和Agilent Bioanalyzer 2100系统评估文库质量。最后,在Illumina NovaSeq平台上对文库进行测序,得到250 bp的成对末端reads。采用统计软件R(R版本R-4.1.2)、Python(Python 3.8.12版本)、Ubuntu(Ubuntu 18.04版本)进行统计分析,当数据非正态分布时,采用面积归一化方法进行正态变换。
代谢物组学
将样品置于EP管中,用井涡法用预冷的80%甲醇重悬。然后将样品在冰上融化并旋转30秒。超声6 min后,在5 000 rpm、4℃离心1 min。将上清液冷冻干燥,用10 %甲醇溶解。最后将溶液注入LC-MS/MS系统分析。UHPLC-MS/MS分析采用Vanquish UHPLC系统(ThermoFisher,德国)与Orbitrap Q ExactiveTM高频质谱仪(ThermoFisher,德国)在Novogene Co.,Ltd(北京,中国)进行。样品以0.2 mL/min的流速以17 min的线性梯度注入到Hypesil金柱(100 × 2.1 mm, 1.9 μm)上。正极性模式的洗脱液为洗脱液A(0.1 % FA in Water)和洗脱液B(Methanol)。负极性模式洗脱液为洗脱液A(5 mM乙酸铵,pH 9.0)和洗脱液B(甲醇)。溶剂梯度设定为:2 % B,1.5 min;2-100% B,3 min;100 % B,10分钟;100-2 % B,10.1 min;Q ExactiveTM高频质谱仪工作在正负极性模式下,喷雾电压3.5 kV,毛细管温度320℃,鞘气流量35 psi,辅气流量10 L/min, S-lens射频电平60,辅气加热器温度350℃。使用Compound Discoverer 3.1(CD3.1, ThermoFisher)对UHPLC-MS/MS生成的原始数据文件进行处理,对每种代谢物进行峰比对、峰拾取和定量。主要参数设置为:保留时间公差0.2 min;实际质量公差,5 ppm;信号强度公差,30 %;信噪比,3;最小强度,等等。之后,将峰值强度归一化为总光谱强度。利用归一化后的数据预测了基于加性离子、分子离子峰和片段离子的分子式。然后与mzCloud (https://www.mzcloud.org/)、mzVault和MassList数据库进行比对,得到准确的定性和相对定量结果。采用统计软件R(R版本R-4.1.2)、Python (Python 3.8.12版本)、Ubuntu(Ubuntu 18.04版本)进行统计分析,当数据非正态分布时,采用面积归一化方法进行正态变换。
超高液相色谱质谱分析
血清样品(10 μl)用40 μl甲醇沉淀,然后在2 000 rpm下涡流3 min,在4℃下在13 000 rpm下离心5 min。然后上清(40 μl)。用甲醇稀释标准原液,制得系列浓度的标准溶液(0.1、0.2、0.5、1、2、5、10、20、50、100、200、300),其中5-羟色胺单位为ng/mL, NE、DA单位为pg/mL。
酶联免疫吸附试验
血清炎症因子IL-6(#MM-0163M2)、IL-1β(#MM-0040M2)和TNF-α (#MM-0132M2)采用ELISA(MEIMIAN,China)根据手册进行检测。用MultiskanTM FC(Thermo Fisher Scientific) 在450 nm处测量吸光度值。根据标准曲线方程计算浓度。
细胞培养
PC12细胞(#CRL-1721™,ATCC)在DMEM(Hyclone, #SH30021.01,Logan,Vtah)中培养,添加10 %胎牛血清(FBS),100 U/mL青霉素和0.1 mg/mL链霉素(#C0222,Beyotime)。细胞保存在加湿培养箱中,空气浓度为95 % /CO2浓度为5 %,温度为37℃。细胞分别用5-HT(5 μM)或ML141(5 mM)处理12 h,再培养24 h。未作其他处理的细胞作为对照。
Cdc42 GTPase活性测定
使用Rac1/Cdc42激活检测试剂盒(#17-441,Sigma-Aldrich)检测Cdc42 GTPase活性。简单地说,用300 µl裂解缓冲液裂解细胞。在离心去除细胞碎片和不溶性物质后,立即将Rac/cdc42 Assay Reagent直接加入裂解液中。在4°C下轻轻摇晃反应混合物60分钟。通过脉冲收集琼脂糖珠(在14000 g微离心机中5秒),并排出上清。用MLB洗三次。琼脂糖珠在适量2×Laemmli样品缓冲液中重悬,煮沸5分钟。10×10 cm微型凝胶每次检测20 µl。收集上清进行SDS-PAGE和随后的免疫印迹分析。用抗cdc 42进行印迹检测,然后用酶标二抗和ECL试剂进行印迹检测。
Western Blotting
PC12细胞用含有1 %磷酸酶和蛋白酶抑制剂(#78442,Thermo Scientific)的冷冻RIPA缓冲液(#89900,Thermo Scientific)匀浆。总蛋白浓度采用BCA测定试剂盒(#P001,Beyotime)测定。通过SDS-PAGE凝胶(#KGP113K,Key GEN Bio Tech)分离蛋白,并通过Trans-Blot®SD细胞(#221BR5621)转移到0.22 μm PVDF膜(#R1JB38277, Millipore)上。用5 % BSA in Tris Buffered Saline with Tween 20 (TBST)在室温下阻断膜1小时,然后与一抗在4℃下孵育过夜。一抗包括CD63 (1:5000,#25682-1-AP,Proteintech)、TSG101(1:2000,#28283-1-AP,,Proteintech)、Cdc42(1:10000,#ab187643,Abcam)、BDNF(1:1000,#ab108319,Abcam)、TrkB(1:1000,#4603,CST)、AKT(1:1000, #AF626,Proteintech)、p-AKT (1:1000,# af8004,CST)、ERK(1:1000, #AF1015,Proteintech)、α-微管蛋白(1:50 000,#66031-1-Ig,Proteintech)。用TBST充分洗涤后,将膜与适当的酶标二抗在室温下孵育1小时。使用ECL检测试剂盒(Millipore,#WBKLS0500)和ChemiDocTM Touch成像系统(Bio-Rad,California, usa)对印迹进行可视化。利用Image lab软件(Bio-Rad, California,U.S.)分析。
免疫荧光染色
用5 % BSA和0.3 % Triton(#XK0193,中国,xuanke,China)在室温下阻断PC12细胞切片1小时,然后用一抗BDNF(1:20 00, #ab187643,Abcam)和NeuN(1:50 00,#12943,CST)染色,PC12细胞切片用BDNF(1:20 00,#ab187643,Abcam)在4°C下染色过夜。然后用1% PBST洗涤切片或切片3次,共15分钟。然后用抗小鼠488(1:80)和抗兔568(1:500)染色1.5小时。在室温下用DAPI(40,60-二氨基-2-苯基吲哚)染色15分钟。在徕卡激光扫描共聚焦显微(德国STELLARIS 5)下观察细胞。
统计分析
所有数值均以平均值±SEM表示。当数据服从正态分布时,采用非配对T检验(假设两个总体具有相同的SD)或Welch校正(不假设SD相等)来检验两组之间的统计显著性。若不存在差异,则采用Mann-Whitney U检验。当数据服从正态分布时,三个或更多组之间的统计显著性采用普通的单向方差分析(ANOVA),然后采用Tukey多重比较检验(假设所有总体具有相同的SD),或Brown-Forsythe和Welch方差检验,然后采用Tamhane T2多重比较检验(不假设SD相等)。若不存在差异,则采用Kruskal-Wallis检验和Dunn多重比较检验进行三组及以上组间的统计学显著性检验。重复测量小鼠体重的统计学显著性评价采用RM双因素方差分析,并采用Tukey多重比较检验(GraphPad Prism 9.3.0)。当p < 0.05时认为差异显著。
引文格式:
Hong, Rui, Lan Luo, Liang Wang, Zhao‐Li Hu, Qi‐Rong Yin, Ming Li, Bin Gu, Bin Wang, Tao Zhuang, Xin‐Yue Zhang, Yuan Zhou, Wan Wang, Lin‐Yan Huang, Bing Gu, and Su‐Hua Qi. 2023. “Lepidium meyenii Walp (Maca)‐derived extracellular vesicles ameliorate depression by promoting 5‐HT synthesis via the modulation of gut–brain axis.” iMeta e116. https://doi.org/10.1002/imt2.116
作者简介

洪蕊(第一作者)
● 徐州医科大学微生物与生化药学,学术硕士。
● 目前研究方向为生化药物研发及相关技术研究,相关学术成果已发表于iMeta、Frontiers in Pharmacology和NEURAL REGENERATION RESEARCH等期刊。

罗兰(第一作者)
● 江苏特聘教授,博士生导师。
● 就职于徐州医科大学医学技术学院,主要从事心脑血管疾病药物筛选和心脏再生研究,以一作/通讯在Circulation Research、Cell Death & Disease、Stem Cell Research & Therapy等期刊发表SCI论文15篇,主持国家自然科学青年1项和面上项目1项、江苏省自然基金青年项目1项。

王亮(第一作者)
● 南方医科大学附属广东省人民医院高层次引进人才,检验科首席研究员,课题组PI,兼任昆士兰大学研究员及博士生导师,西澳大学研究员及伊迪斯·科文大学客座教授。
● 本科毕业于中南大学湘雅医学院,博士毕业于西澳大学病理与医学检验学院,加拿大康考迪亚大学及澳大利亚科廷大学博士后。入选江苏省双创计划,江苏省青蓝工程,江苏省“第十三批科技镇长团”。Frontiers in Microbiology副主编,BMC Microbiology及PeerJ等SCI期刊编委,iMeta青年编委。主要从事计算和实验联合的生物医学研究,主持国自然基金2项,省自然基金2项,参与科技部重大项目4项,发表英文论文100余篇(通讯/一作SCI论文57篇),主编《系统生物学》及《精油的科学》等中英文教材专著6部。

胡兆丽(第一作者)
● 徐州医科大学生物化学教研室&江苏省脑病生物信息重点实验室。
● 任职于徐州医科大学生物化学教研室&江苏省脑病生物信息重点实验室主要从事神经退行性疾病分子机制的研究工作。曾参与多项国家自然科学基金项目,主持徐州市自然科技计划项目一项。以第一作者先后在Brain, behavior, and immunity、Stem cell research & therapy杂志发表文章。

齐素华(通讯作者)
● 博士,教授,博士生导师,徐州医科大学医学技术学院 实验室主任、徐州市临床检验诊断学重点实验室主任。
● 主要从事临床检验诊断学技术与心脑血管发病机制的研究,任江苏省神经科学学会委员、江苏省卒中学会委员、江苏省神经免疫学会委员等。获江苏省“333”工程高层次人才、江苏省“六大人才高峰”培养对象、徐州市优秀科技工作者等称号。曾获中华医学科技奖二等奖1项、江苏省医学科技奖二等奖1项、三等奖1项。先后在J. Am. Chem, Neuropathology & Applied Neurobiology,Neuropharmacology 等国际著名期刊,以第一作者或通讯作者发表论文SCI论文50余篇,获批发明专利6项。

顾兵(通讯作者)
● 医学博士、教授、博士生导师,广东省人民医院检验科主任,广东省临床基因检测质量控制中心主任/广东省人民医院检验科主任美国普渡大学及UCLA访问学者,国家重点研发计划首席科学家、广东省“珠江人才”领军人才、省“科教强卫”医学重点人才。
● 任中华医学会检验分会青委会副主委、中国医学装备协会检验医学分会副会长、广东省临床基因检测质控中心主任、广东省卫生经济学会检验经济分会会长、J Lab Precis Med执行主编。从事重大传染病快速检测新技术与防控研究,主持国家重点研发计划1项、国家自然科学基金5项、省部级课题8项。以第一或通迅作者在Emerg Infect Dis、Emerg Microbes Infect、J Clin Microbiol等期刊发表SCI论文114篇,10分以上13篇,累计影响因子664分;主编及副主编专著18部;获授权专利9项。
更多推荐
(▼ 点击跳转)
iMeta | 德国国家肿瘤中心顾祖光发表复杂热图(ComplexHeatmap)可视化方法
iMeta | 浙大倪艳组MetOrigin实现代谢物溯源和肠道微生物组与代谢组整合分析


1卷1期

1卷2期

1卷3期

1卷4期

2卷1期
期刊简介
“iMeta” 是由威立、肠菌分会和本领域数百位华人科学家合作出版的开放获取期刊,主编由中科院微生物所刘双江研究员和荷兰格罗宁根大学傅静远教授担任。目的是发表原创研究、方法和综述以促进宏基因组学、微生物组和生物信息学发展。目标是发表前10%(IF > 15)的高影响力论文。期刊特色包括视频投稿、可重复分析、图片打磨、青年编委、前3年免出版费、50万用户的社交媒体宣传等。2022年2月正式创刊发行!
联系我们
iMeta主页:http://www.imeta.science
出版社:https://onlinelibrary.wiley.com/journal/2770596x
投稿:https://mc.manuscriptcentral.com/imeta
邮箱:[email protected]
直接关联